

MicroRNA-570 is a novel regulator of cellular senescence and inflammaging
- PMID: 30156909
- PMCID: PMC6338629
- DOI: 10.1096/fj.201800965R
MicroRNA-570 is a novel regulator of cellular senescence and inflammaging
Abstract
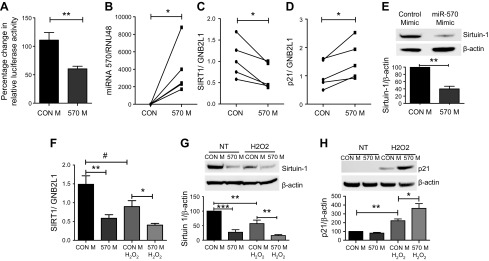
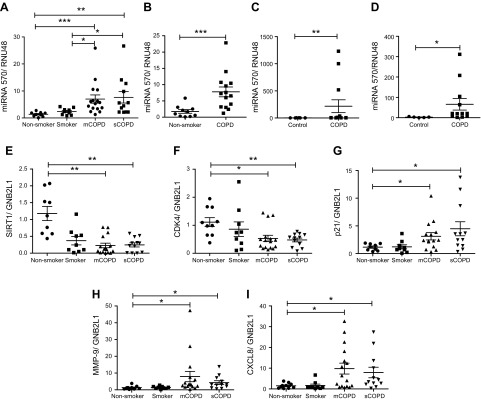
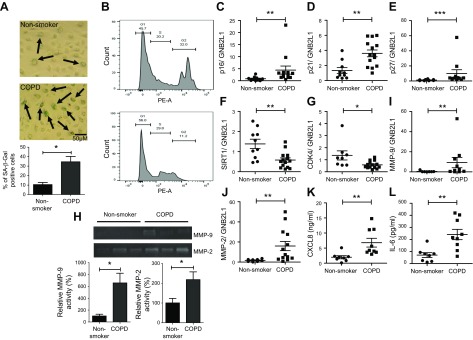
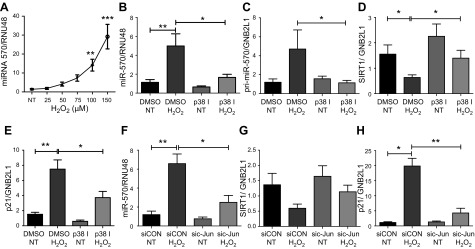
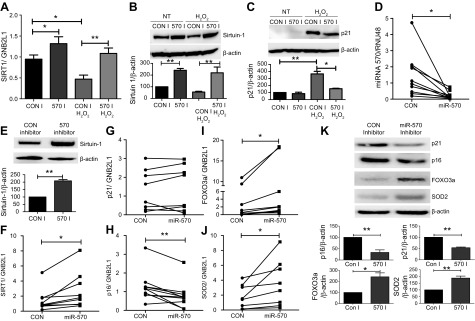
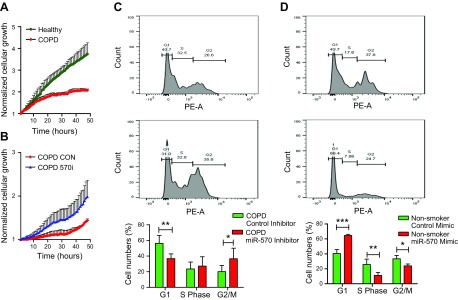
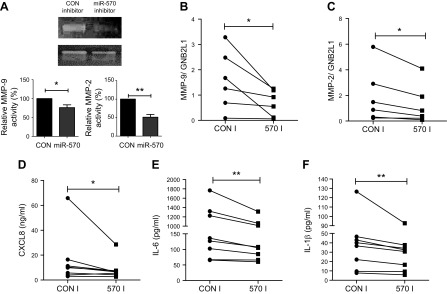
Diseases of accelerated aging often occur together (multimorbidity), and their prevalence is increasing, with high societal and health care costs. Chronic obstructive pulmonary disease (COPD) is one such condition, in which one half of patients exhibit ≥4 age-related diseases. Diseases of accelerated aging share common molecular pathways, which lead to the detrimental accumulation of senescent cells. These senescent cells no longer divide but release multiple inflammatory proteins, known as the senescence-associated secretory phenotype, which may perpetuate and speed disease. Here, we show that inhibiting miR-570-3p, which is increased in COPD cells, reverses cellular senescence by restoring the antiaging molecule sirtuin-1. MiR-570-3p is induced by oxidative stress in airway epithelial cells through p38 MAP kinase-c-Jun signaling and drives senescence by inhibiting sirtuin-1. Inhibition of elevated miR-570-3p in COPD small airway epithelial cells, using an antagomir, restores sirtuin-1 and suppresses markers of cellular senescence (p16INK4a, p21Waf1, and p27Kip1), thereby restoring cellular growth by allowing progression through the cell cycle. MiR-570-3p inhibition also suppresses the senescence-associated secretory phenotype (matrix metalloproteinases-2/9, C-X-C motif chemokine ligand 8, IL-1β, and IL-6). Collectively, these data suggest that inhibiting miR-570-3p rejuvenates cells via restoration of sirtuin-1, reducing many of the abnormalities associated with cellular senescence.-Baker, J. R., Vuppusetty, C., Colley, T., Hassibi, S., Fenwick, P. S., Donnelly, L. E., Ito, K., Barnes, P. J. MicroRNA-570 is a novel regulator of cellular senescence and inflammaging.
Keywords: COPD; cell cycle; epithelial cells; inflammation; miRNA.
Conflict of interest statement
The authors thank Prof. Jim Hogg (University of British Columbia, Vancouver, BC, Canada) for kindly providing peripheral lung tissue samples and Dr. Andriana I. Papaioannou (Sismanogleio Hospital, Athens, Greece) for providing the sputum samples. This work was funded by the British Lung Foundation Grant (JFRG17-7), Wellcome Trust Programme Grant (093080/Z/10/Z), and supported by the NIHR Respiratory Disease Biomedical Research Unit at the Royal Brompton and Harefield NHS Foundation Trust and Imperial College London. The authors declare no conflicts of interest.
Figures
References
-
- Lozano, R., Naghavi, M., Foreman, K., Lim, S., Shibuya, K., Aboyans, V., Abraham, J., Adair, T., Aggarwal, R., Ahn, S. Y., Alvarado, M., Anderson, H. R., Anderson, L. M., Andrews, K. G., Atkinson, C., Baddour, L. M., Barker-Collo, S., Bartels, D. H., Bell, M. L., Benjamin, E. J., Bennett, D., Bhalla, K., Bikbov, B., Bin Abdulhak, A., Birbeck, G., Blyth, F., Bolliger, I., Boufous, S., Bucello, C., Burch, M., Burney, P., Carapetis, J., Chen, H., Chou, D., Chugh, S. S., Coffeng, L. E., Colan, S. D., Colquhoun, S., Colson, K. E., Condon, J., Connor, M. D., Cooper, L. T., Corriere, M., Cortinovis, M., de Vaccaro, K. C., Couser, W., Cowie, B. C., Criqui, M. H., Cross, M., Dabhadkar, K. C., Dahodwala, N., De Leo, D., Degenhardt, L., Delossantos, A., Denenberg, J., Des Jarlais, D. C., Dharmaratne, S. D., Dorsey, E. R., Driscoll, T., Duber, H., Ebel, B., Erwin, P. J., Espindola, P., Ezzati, M., Feigin, V., Flaxman, A. D., Forouzanfar, M. H., Fowkes, F. G., Franklin, R., Fransen, M., Freeman, M. K., Gabriel, S. E., Gakidou, E., Gaspari, F., Gillum, R. F., Gonzalez-Medina, D., Halasa, Y. A., Haring, D., Harrison, J. E., Havmoeller, R., Hay, R. J., Hoen, B., Hotez, P. J., Hoy, D., Jacobsen, K. H., James, S. L., Jasrasaria, R., Jayaraman, S., Johns, N., Karthikeyan, G., Kassebaum, N., Keren, A., Khoo, J. P., Knowlton, L. M., Kobusingye, O., Koranteng, A., Krishnamurthi, R., Lipnick, M., Lipshultz, S. E., Ohno, S. L., Mabweijano, J., MacIntyre, M. F., Mallinger, L., March, L., Marks, G. B., Marks, R., Matsumori, A., Matzopoulos, R., Mayosi, B. M., McAnulty, J. H., McDermott, M. M., McGrath, J., Mensah, G. A., Merriman, T. R., Michaud, C., Miller, M., Miller, T. R., Mock, C., Mocumbi, A. O., Mokdad, A. A., Moran, A., Mulholland, K., Nair, M. N., Naldi, L., Narayan, K. M., Nasseri, K., Norman, P., O’Donnell, M., Omer, S. B., Ortblad, K., Osborne, R., Ozgediz, D., Pahari, B., Pandian, J. D., Rivero, A. P., Padilla, R. P., Perez-Ruiz, F., Perico, N., Phillips, D., Pierce, K., Pope C. A. III, Porrini, E., Pourmalek, F., Raju, M., Ranganathan, D., Rehm, J. T., Rein, D. B., Remuzzi, G., Rivara, F. P., Roberts, T., De León, F. R., Rosenfeld, L. C., Rushton, L., Sacco, R. L., Salomon, J. A., Sampson, U., Sanman, E., Schwebel, D. C., Segui-Gomez, M., Shepard, D. S., Singh, D., Singleton, J., Sliwa, K., Smith, E., Steer, A., Taylor, J. A., Thomas, B., Tleyjeh, I. M., Towbin, J. A., Truelsen, T., Undurraga, E. A., Venketasubramanian, N., Vijayakumar, L., Vos, T., Wagner, G. R., Wang, M., Wang, W., Watt, K., Weinstock, M. A., Weintraub, R., Wilkinson, J. D., Woolf, A. D., Wulf, S., Yeh, P. H., Yip, P., Zabetian, A., Zheng, Z. J., Lopez, A. D., Murray, C. J., AlMazroa, M. A., Memish, Z. A. (2012) Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010 [published erratum appears in Lancet 2013;381:628] . Lancet 380, 2095–2128 10.1016/S0140-6736(12)61728-0 - DOI - PMC - PubMed
-
- Vanfleteren, L. E., Spruit, M. A., Groenen, M., Gaffron, S., van Empel, V. P., Bruijnzeel, P. L., Rutten, E. P., Op ’t Roodt, J., Wouters, E. F., Franssen, F. M. (2013) Clusters of comorbidities based on validated objective measurements and systemic inflammation in patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 187, 728–735 10.1164/rccm.201209-1665OC - DOI - PubMed
-
- Kennedy, B. K., Berger, S. L., Brunet, A., Campisi, J., Cuervo, A. M., Epel, E. S., Franceschi, C., Lithgow, G. J., Morimoto, R. I., Pessin, J. E., Rando, T. A., Richardson, A., Schadt, E. E., Wyss-Coray, T., Sierra, F. (2014) Geroscience: linking aging to chronic disease. Cell 159, 709–713 10.1016/j.cell.2014.10.039 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
