

Engineering plant architecture via CRISPR/Cas9-mediated alteration of strigolactone biosynthesis
- PMID: 30157762
- PMCID: PMC6116466
- DOI: 10.1186/s12870-018-1387-1
Engineering plant architecture via CRISPR/Cas9-mediated alteration of strigolactone biosynthesis
Abstract
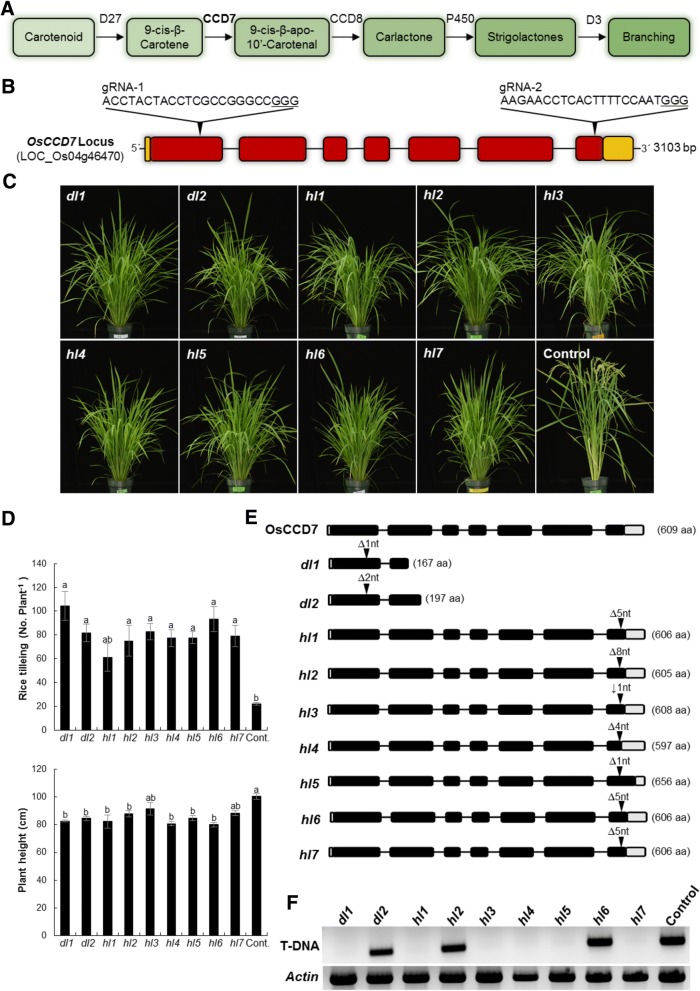
Background: Precision plant genome engineering holds much promise for targeted improvement of crop traits via unprecedented single-base level control over the genetic material. Strigolactones (SLs) are a key determinant of plant architecture, known for their role in inhibiting shoot branching (tillering).
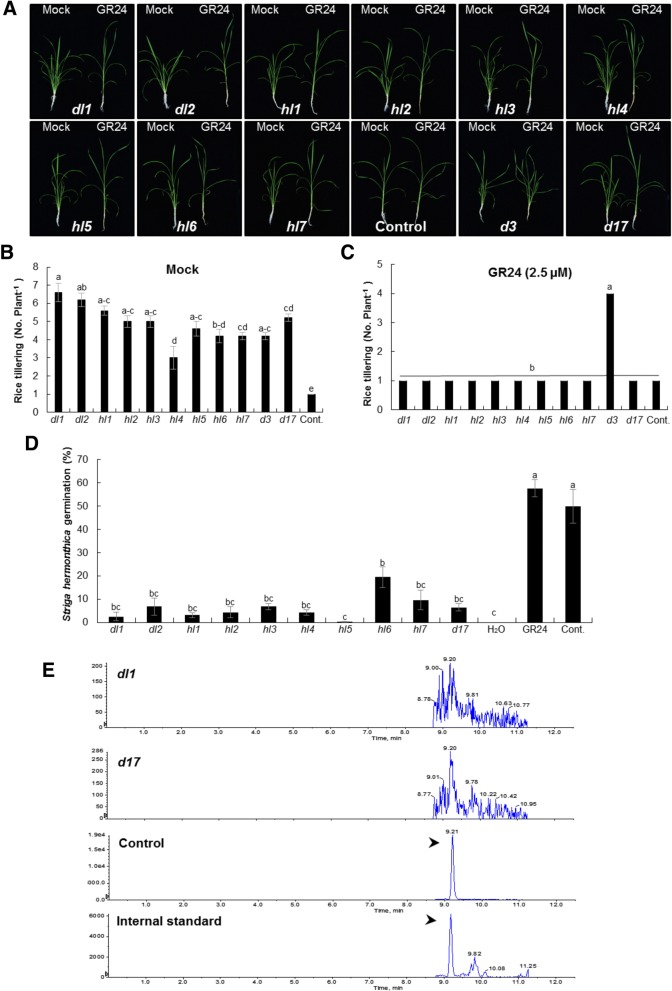
Results: We used CRISPR/Cas9 in rice (Oryza sativa) for targeted disruption of CAROTENOID CLEAVAGE DIOXYGENASE 7 (CCD7), which controls a key step in SL biosynthesis. The ccd7 mutants exhibited a striking increase in tillering, combined with a reduced height, which could be rescued by application of the synthetic SL analog GR24. Striga germination assays and liquid chromatography-mass spectrometry analysis showed that root exudates of ccd7 mutants were also SL deficient.
Conclusions: Taken together, our results show the potential and feasibility of the use of the CRISPR/Cas9 system for targeted engineering of plant architecture and for elucidating the molecular underpinnings of architecture-related traits.
Keywords: CCD7; CRISPR/Cas9; Carotenoid cleavage dioxygenases; Carotenoids; Crop improvement; Genome editing; Plant architecture; Rice engineering; Strigolactones.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Abe S, Sado A, Tanaka K, Kisugi T, Asami K, Ota S, Kim HI, Yoneyama K, Xie X, Ohnishi T, Seto Y, Yamaguchi S, Akiyama K, Yoneyama K, Nomura T. Carlactone is converted to carlactonoic acid by MAX1 in Arabidopsis and its methyl ester can directly interact with AtD14 in vitro. Proc Natl Acad Sci U S A. 2014;111(50):18084–18089. doi: 10.1073/pnas.1410801111. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
