

Short term optical defocus perturbs normal developmental shifts in retina/RPE protein abundance
- PMID: 30157773
- PMCID: PMC6116556
- DOI: 10.1186/s12861-018-0177-1
Short term optical defocus perturbs normal developmental shifts in retina/RPE protein abundance
Abstract
Background: Myopia (short-sightedness) affects approximately 1.4 billion people worldwide, and prevalence is increasing. Animal models induced by defocusing lenses show striking similarity with human myopia in terms of morphology and the implicated genetic pathways. Less is known about proteome changes in animals. Thus, the present study aimed to improve understanding of protein pathway responses to lens defocus, with an emphasis on relating expression changes to no lens control development and identifying bidirectional and/or distinct pathways across myopia and hyperopia (long-sightedness) models.
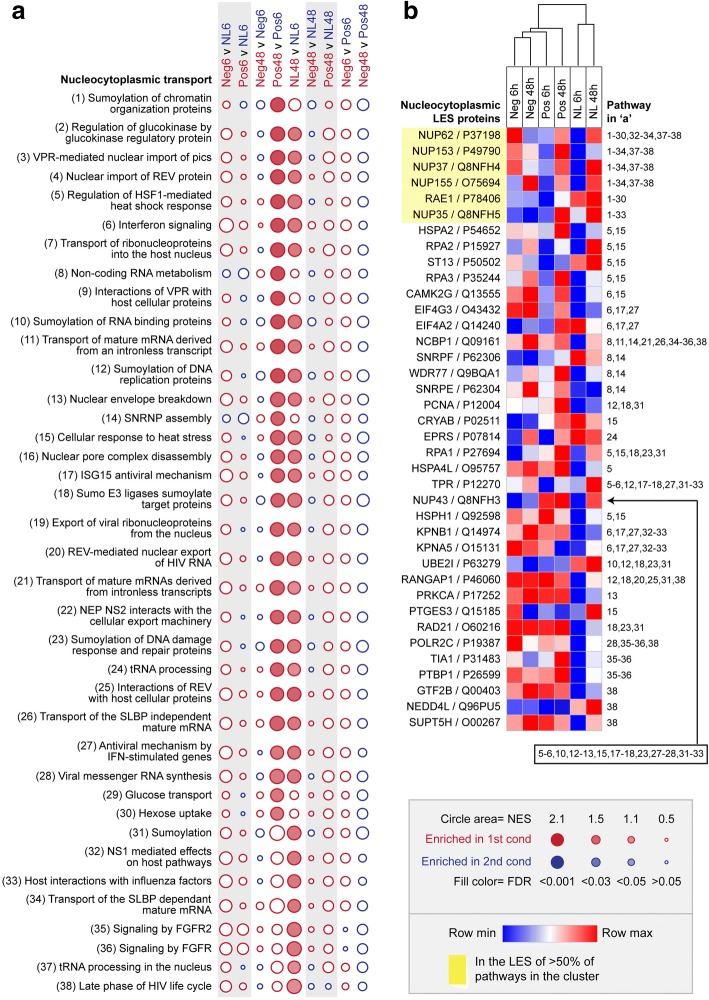
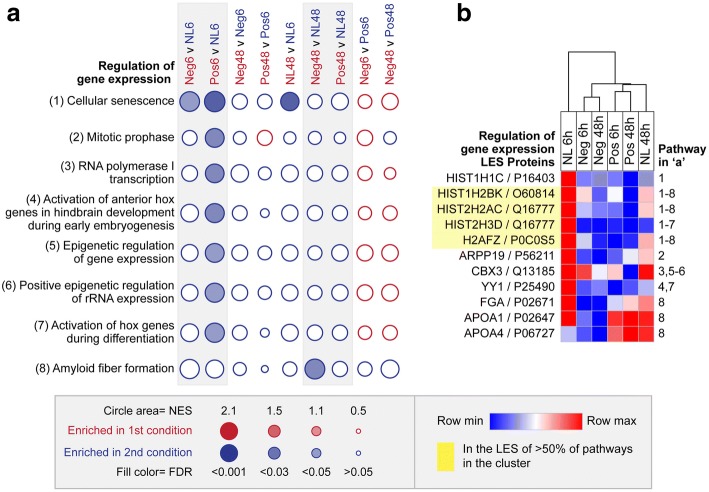
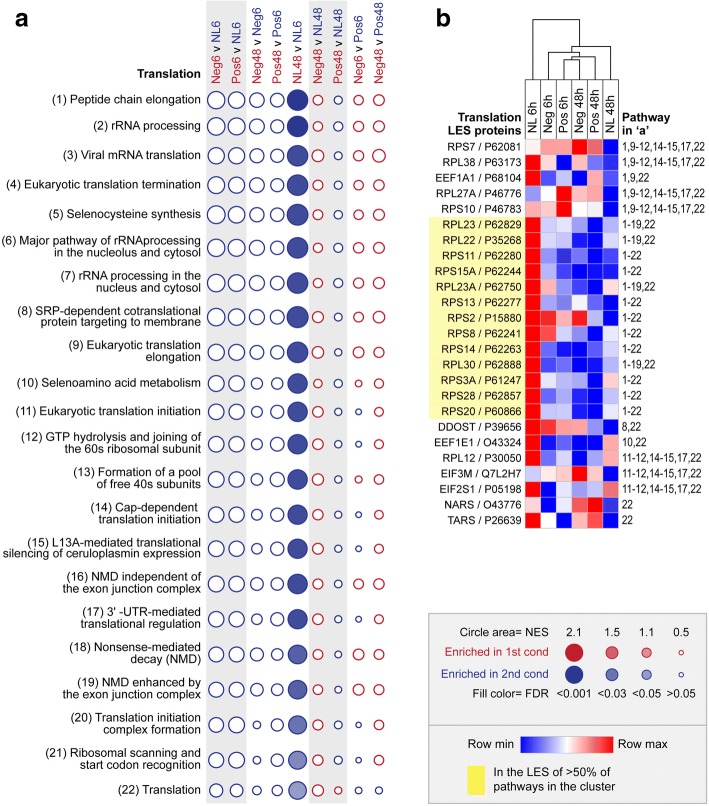
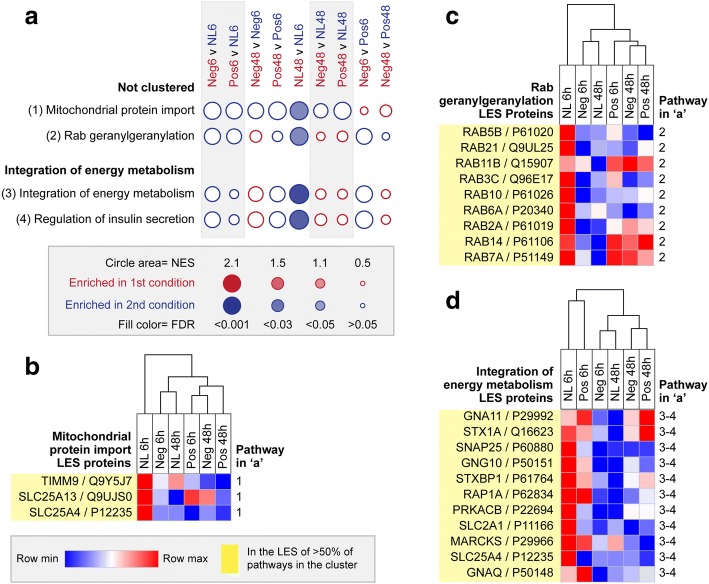
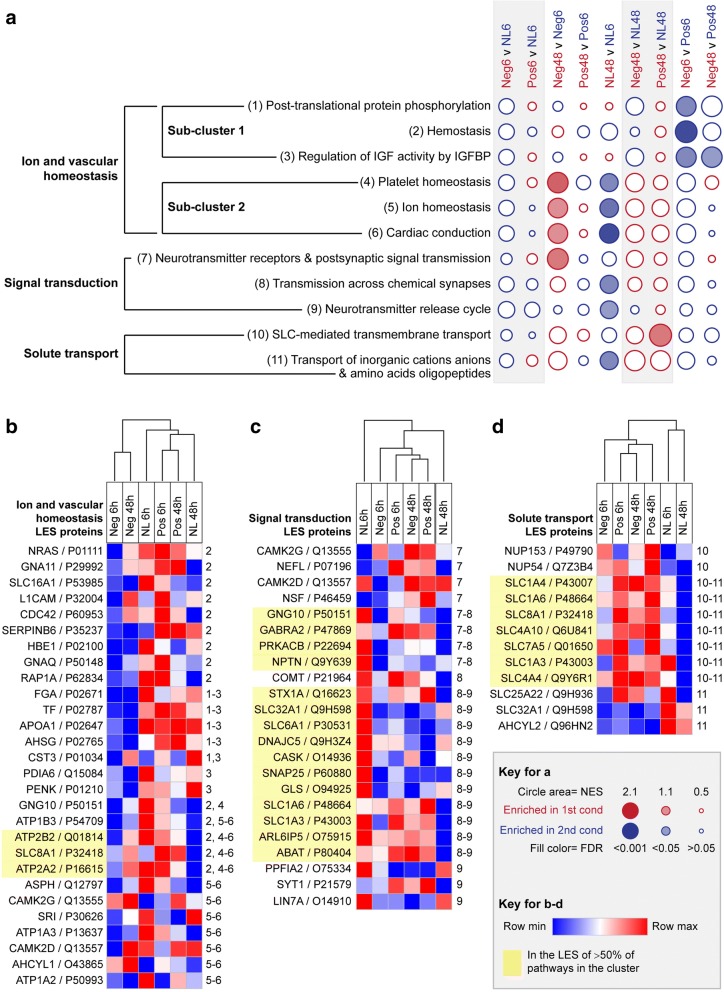
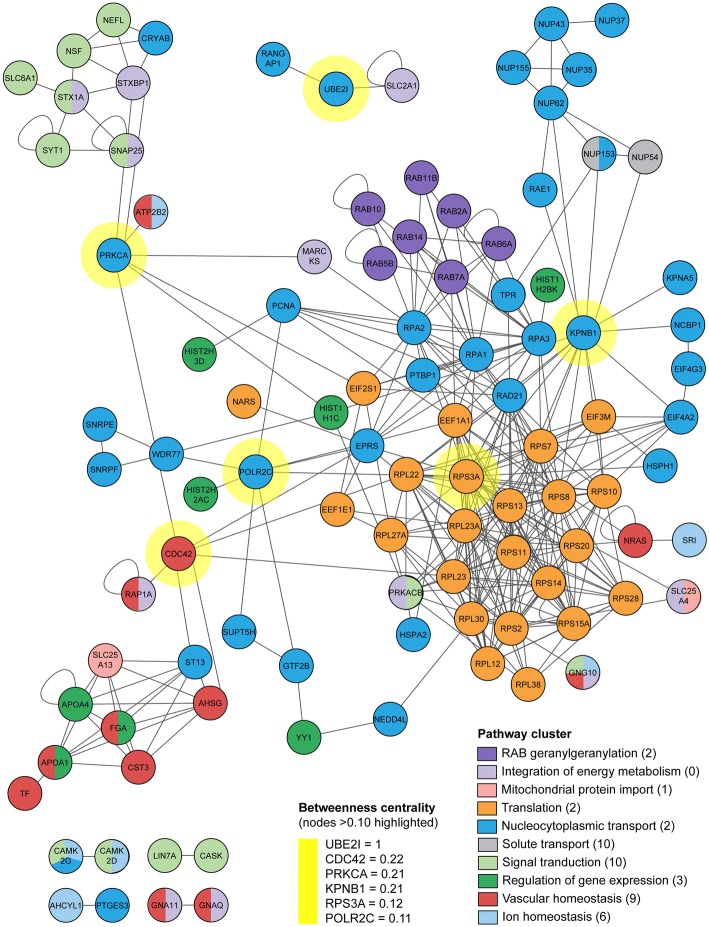
Results: Quantitative label-free proteomics and gene set enrichment analysis (GSEA) were used to examine protein pathway expression in the retina/RPE of chicks following 6 h and 48 h of myopia induction with - 10 dioptre (D) lenses, hyperopia induction with +10D lenses, or normal no lens rearing. Seventy-one pathways linked to cell development and neuronal maturation were differentially enriched between 6 and 48 h in no lens chicks. The majority of these normal developmental changes were disrupted by lens-wear (47 of 71 pathways), however, only 11 pathways displayed distinct expression profiles across the lens conditions. Most notably, negative lens-wear induced up-regulation of proteins involved in ATP-driven ion transport, calcium homeostasis, and GABA signalling between 6 and 48 h, while the same proteins were down-regulated over time in normally developing chicks. Glutamate and bicarbonate/chloride transporters were also down-regulated over time in normally developing chicks, and positive lens-wear inhibited this down-regulation.
Conclusions: The chick retina/RPE proteome undergoes extensive pathway expression shifts during normal development. Most of these pathways are further disrupted by lens-wear. The identified expression patterns suggest close interactions between neurotransmission (as exemplified by increased GABA receptor and synaptic protein expression), cellular ion homeostasis, and associated energy resources during myopia induction. We have also provided novel evidence for changes to SLC-mediated transmembrane transport during hyperopia induction, with potential implications for signalling at the photoreceptor-bipolar synapse. These findings reflect a key role for perturbed neurotransmission and ionic homeostasis in optically-induced refractive errors, and are predicted by our Retinal Ion Driven Efflux (RIDE) model.
Keywords: Chick; Development; Emmetropization; Hyperopia; Mass spectrometry; Myopia; Neurotransmission; Proteomics.
Conflict of interest statement
Ethics approval
The protocols used in this study were approved by the La Trobe University Animal Ethics Committee under application number AEC14–60.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
