

One month of hyperglycemia alters spectral responses of the zebrafish photopic electroretinogram
- PMID: 30158110
- PMCID: PMC6215424
- DOI: 10.1242/dmm.035220
One month of hyperglycemia alters spectral responses of the zebrafish photopic electroretinogram
Abstract
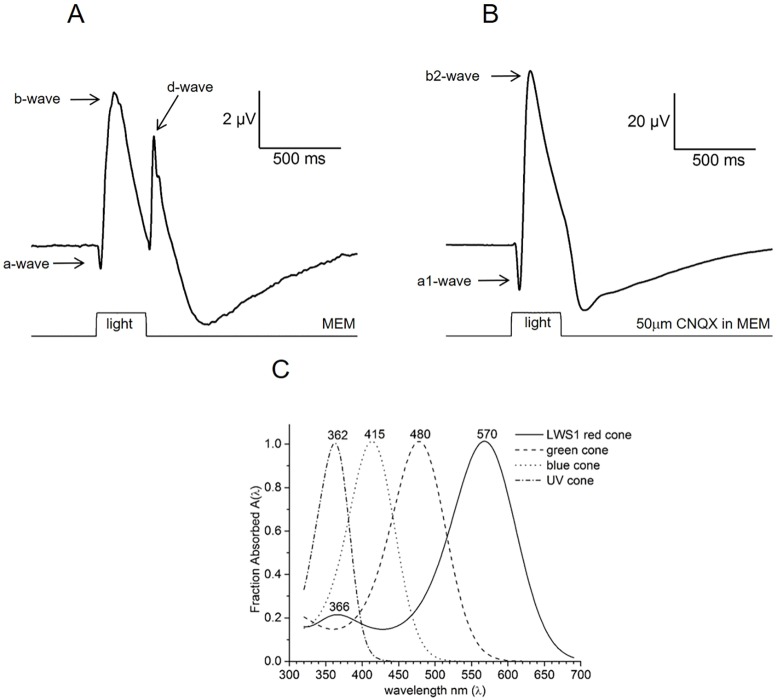
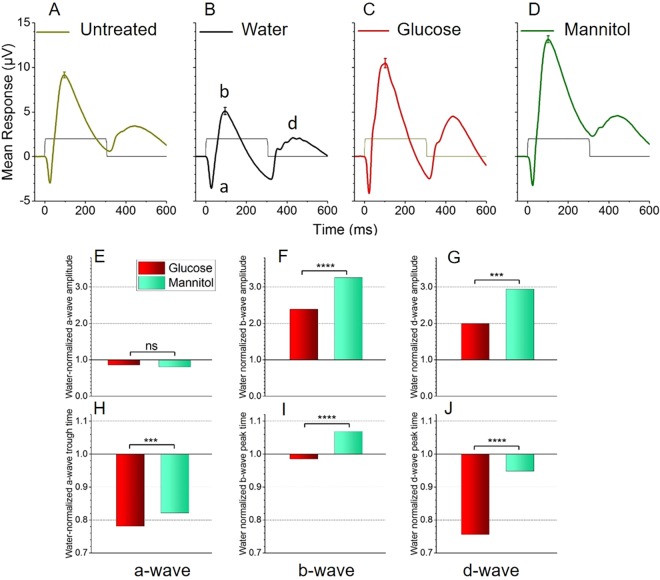
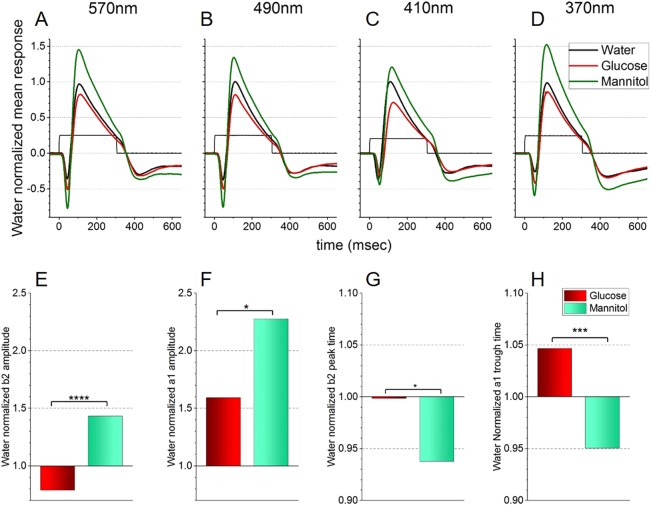
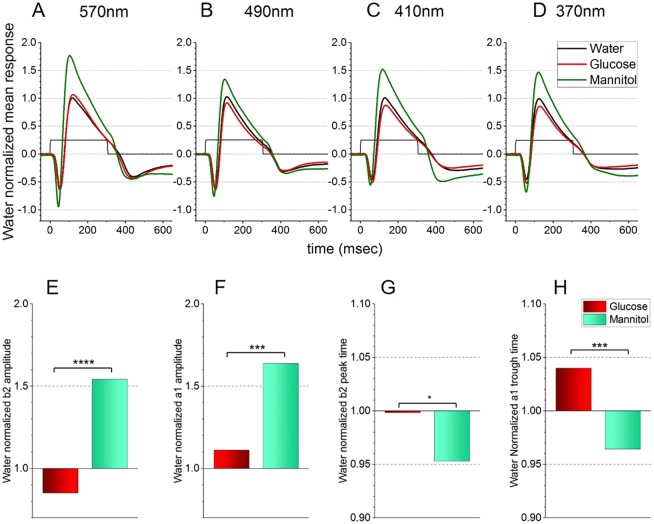
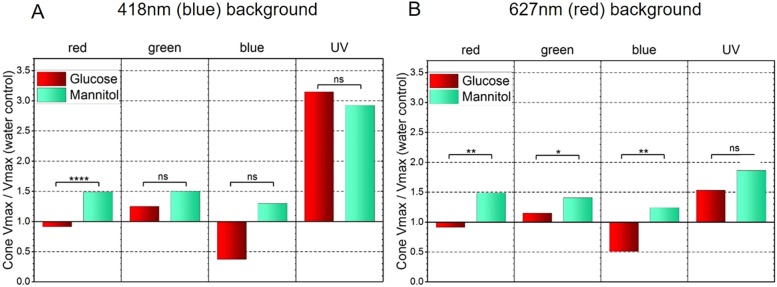
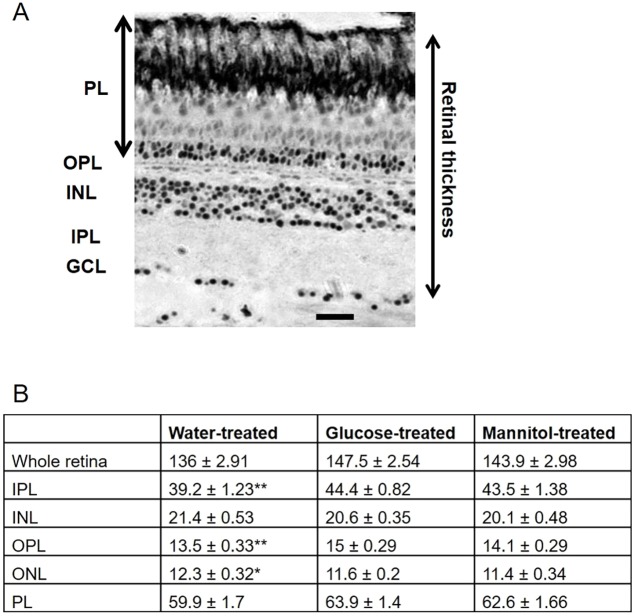
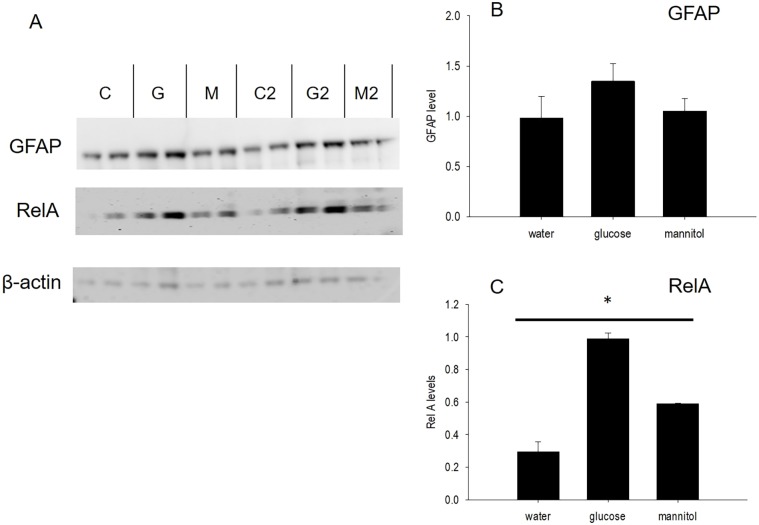
Prolonged hyperglycemia can alter retinal function, ultimately resulting in blindness. Adult zebrafish adults exposed to alternating conditions of 2% glucose/0% glucose display a 3× increase in blood sugar levels. After 4 weeks of treatment, electroretinograms (ERGs) were recorded from isolated, perfused, in vitro eyecups. Control animals were exposed to alternating 2% mannitol/0% mannitol (osmotic control) or to alternating water (0% glucose/0% glucose; handling control). Two types of ERGs were recorded: (1) native ERGs measured using white-light stimuli and medium without synaptic blockers; and (2) spectral ERGs measured with an AMPA/kainate receptor antagonist, isolating photoreceptor-to-ON-bipolar-cell synapses, and a spectral protocol that separated red (R), green (G), blue (B) and UV cone signals. Retinas were evaluated for changes in layer thickness and for the inflammatory markers GFAP and Nf-κB (RelA or p65). In native ERGs, hyperglycemic b- and d-waves were lower in amplitude than the b- and d-waves of mannitol controls. Alteration of waveshape became severe, with b-waves becoming more transient and ERG responses showing more PIII-like (a-wave) characteristics. For spectral ERGs, waveshape appeared similar in all treatment groups. However, a1- and b2-wave implicit times were significantly longer, and amplitudes were significantly reduced, in response to hyperglycemic treatment, owing to the functional reduction in signals from R, G and B cones. Nf-κB increased significantly in hyperglycemic retinas, but the increase in GFAP was not significant and retinal layer thickness was unaffected. Thus, prolonged hyperglycemia triggers an inflammatory response and functional deficits localized to specific cone types, indicating the rapid onset of neural complications in the zebrafish model of diabetic retinopathy.
Keywords: A-wave; B-wave; Diabetes; Glucose; ON bipolar; Outer retina.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Acerete L., Balasch J. C., Espinosa E., Josa A. and Tort L. (2004). Physiological responses in Eurasian perch (Perca fluviatilis, L.) subjected to stress by transport and handling. Aquaculture 237, 167-178. 10.1016/j.aquaculture.2004.03.018 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
