

The Drosophila Epidermal Growth Factor Receptor does not act in the nucleus
- PMID: 30158176
- PMCID: PMC6176925
- DOI: 10.1242/jcs.220251
The Drosophila Epidermal Growth Factor Receptor does not act in the nucleus
Abstract
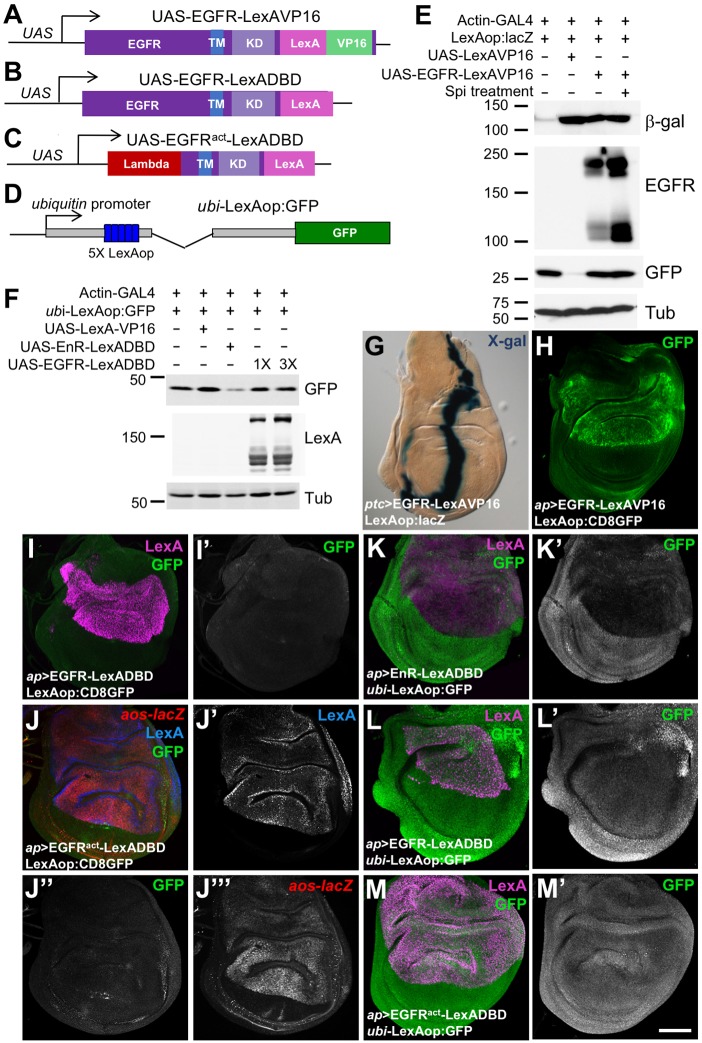
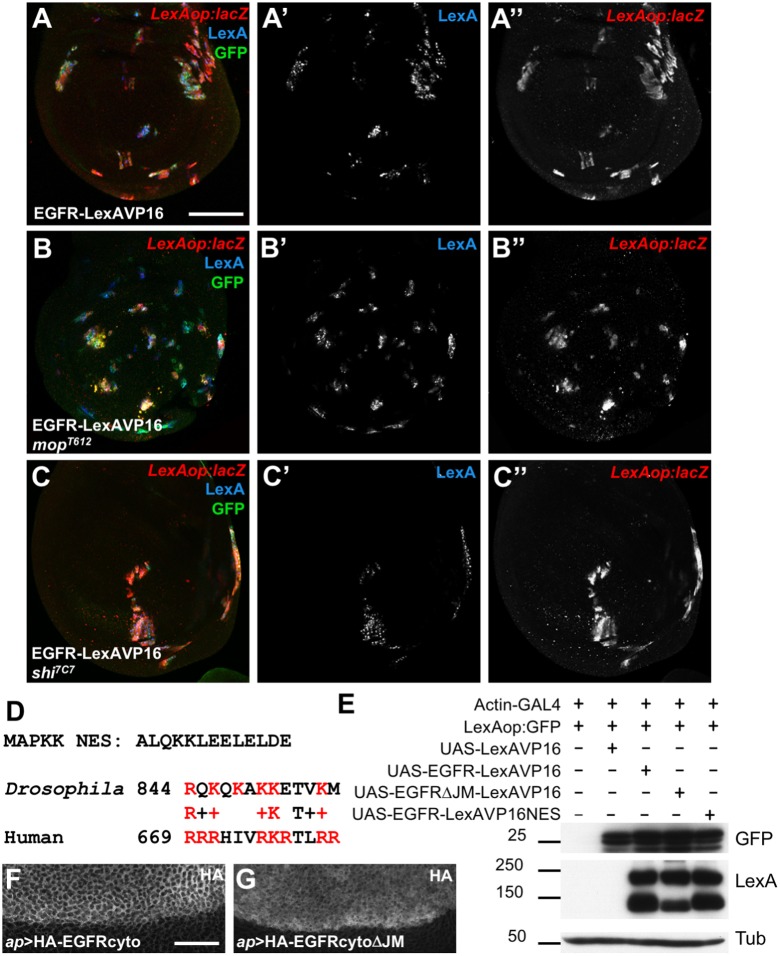
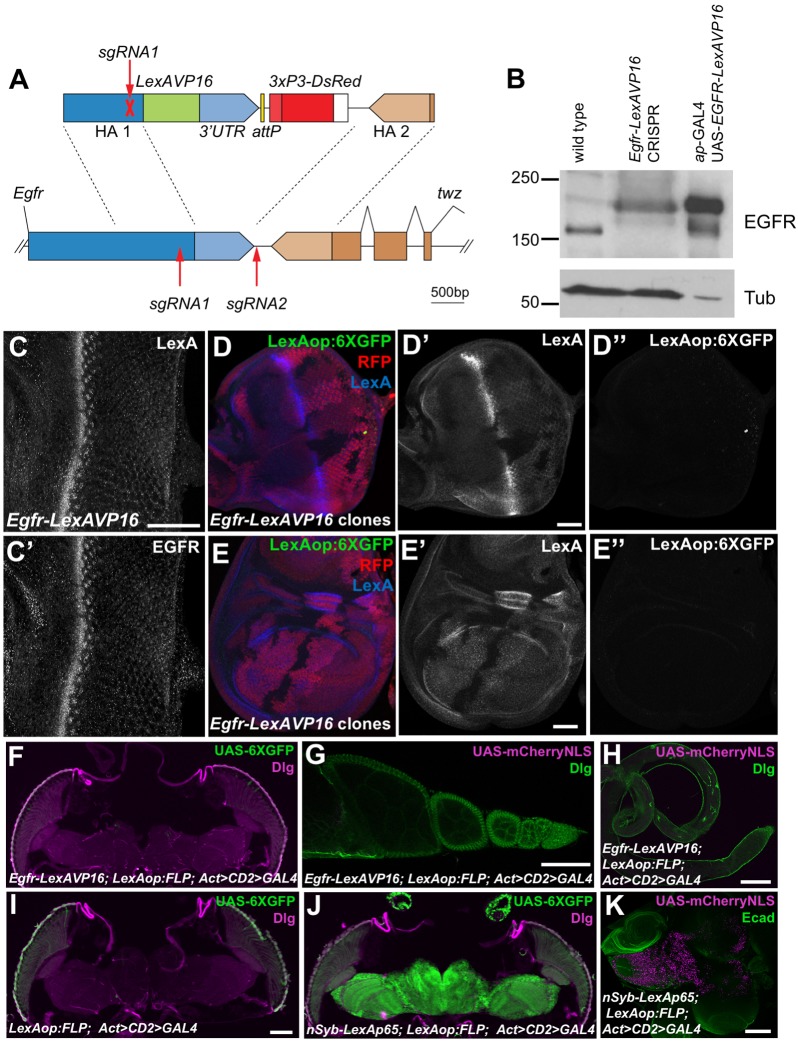
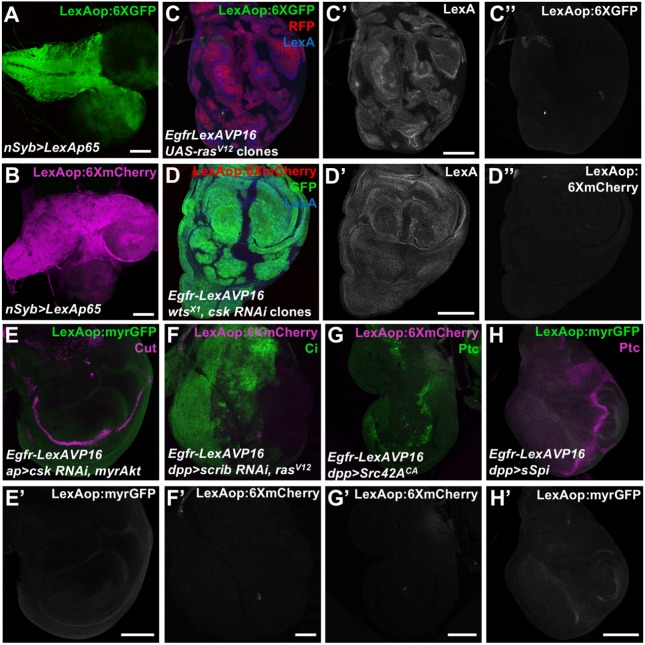
Mammalian members of the ErbB family, including the epidermal growth factor receptor (EGFR), can regulate transcription, DNA replication and repair through nuclear entry of either the full-length proteins or their cleaved cytoplasmic domains. In cancer cells, these nuclear functions contribute to tumor progression and drug resistance. Here, we examined whether the single Drosophila EGFR can also localize to the nucleus. A chimeric EGFR protein fused at its cytoplasmic C-terminus to DNA-binding and transcriptional activation domains strongly activated transcriptional reporters when overexpressed in cultured cells or in vivo However, this activity was independent of cleavage and endocytosis. Without an exogenous activation domain, EGFR fused to a DNA-binding domain did not activate or repress transcription. Addition of the same DNA-binding and transcriptional activation domains to the endogenous Egfr locus through genome editing led to no detectable reporter expression in wild-type or oncogenic contexts. These results show that, when expressed at physiological levels, the cytoplasmic domain of the Drosophila EGFR does not have access to the nucleus. Therefore, nuclear EGFR functions are likely to have evolved after vertebrates and invertebrates diverged.
Keywords: Cancer; EGFR; Nucleus; Transcription.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Beguelin W., Diaz Flaque M. C., Proietti C. J., Cayrol F., Rivas M. A., Tkach M., Rosemblit C., Tocci J. M., Charreau E. H., Schillaci R. et al. (2010). Progesterone receptor induces ErbB-2 nuclear translocation to promote breast cancer growth via a novel transcriptional effect: ErbB-2 function as a coactivator of Stat3. Mol. Cell. Biol. 30, 5456-5472. 10.1128/MCB.00012-10 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
