

Cotranslational assembly of protein complexes in eukaryotes revealed by ribosome profiling
- PMID: 30158700
- PMCID: PMC6372068
- DOI: 10.1038/s41586-018-0462-y
Cotranslational assembly of protein complexes in eukaryotes revealed by ribosome profiling
Erratum in
-
Author Correction: Cotranslational assembly of protein complexes in eukaryotes revealed by ribosome profiling.Nature. 2026 Feb;650(8103):E23. doi: 10.1038/s41586-026-10200-8. Nature. 2026. PMID: 41639466 No abstract available.
Abstract
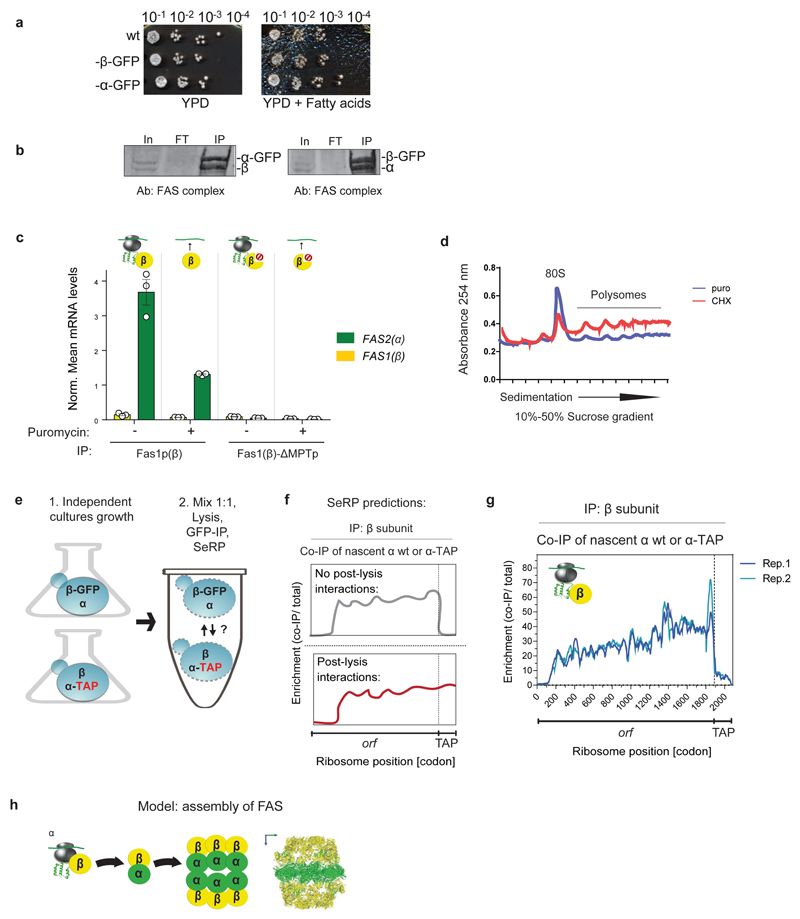
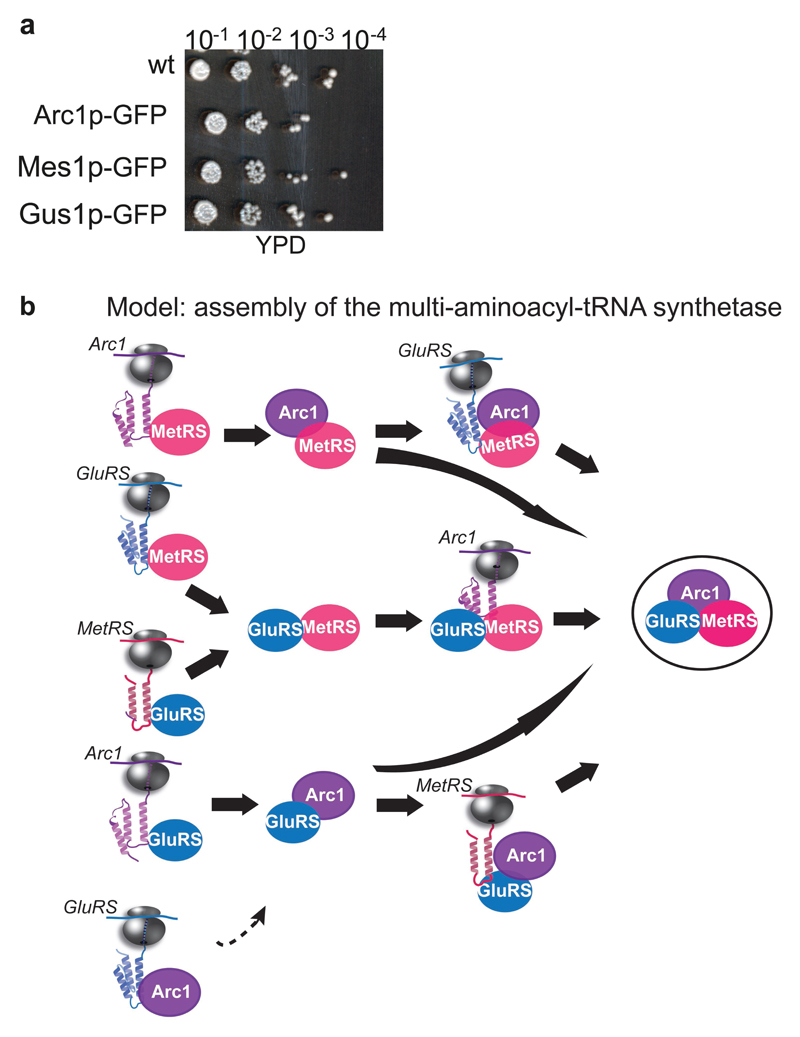
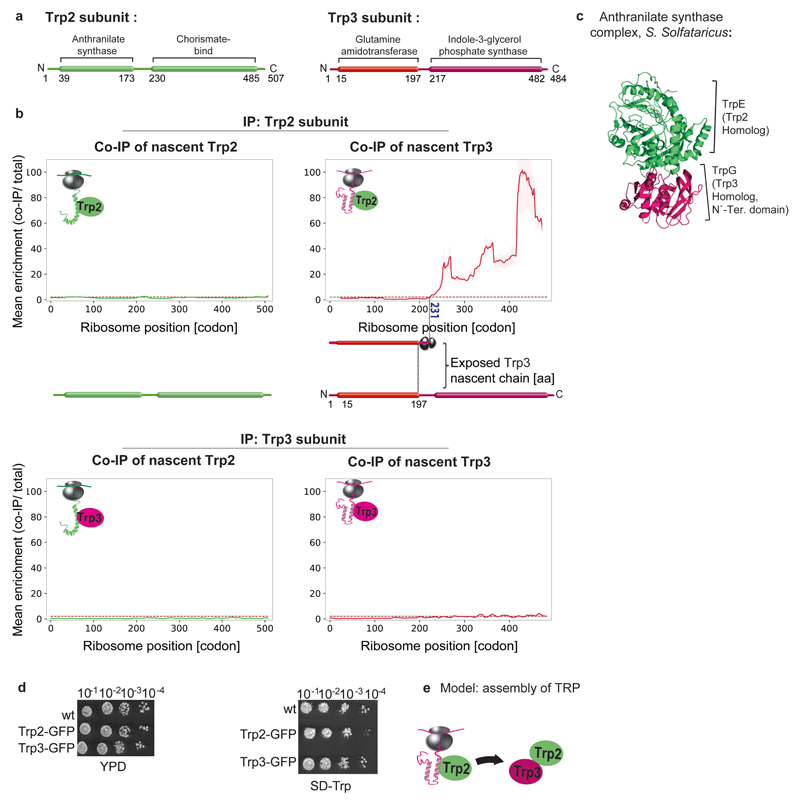
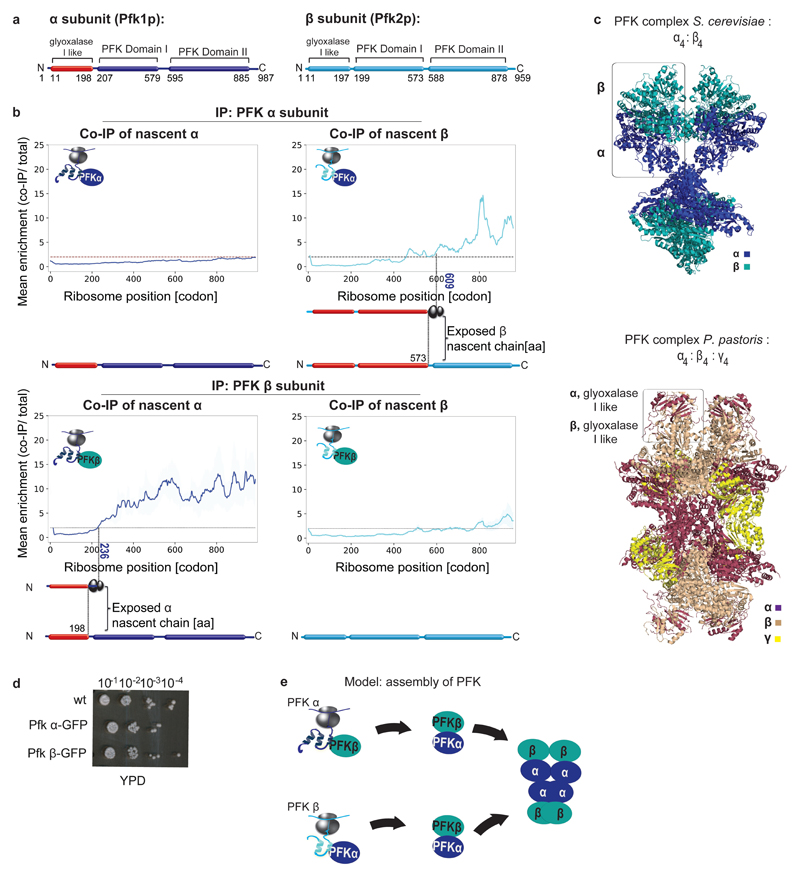
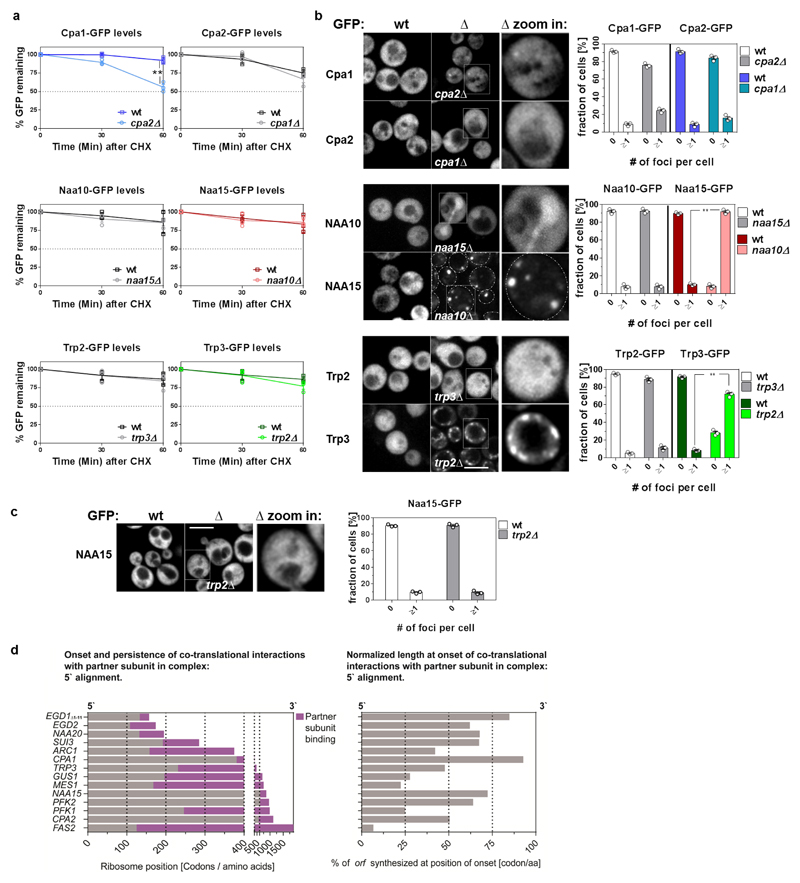
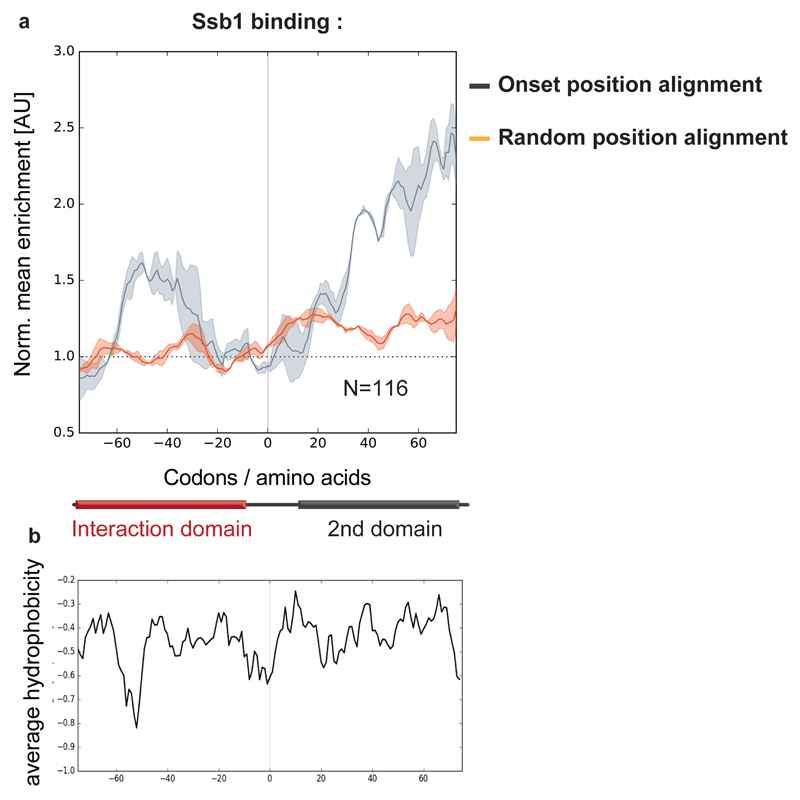
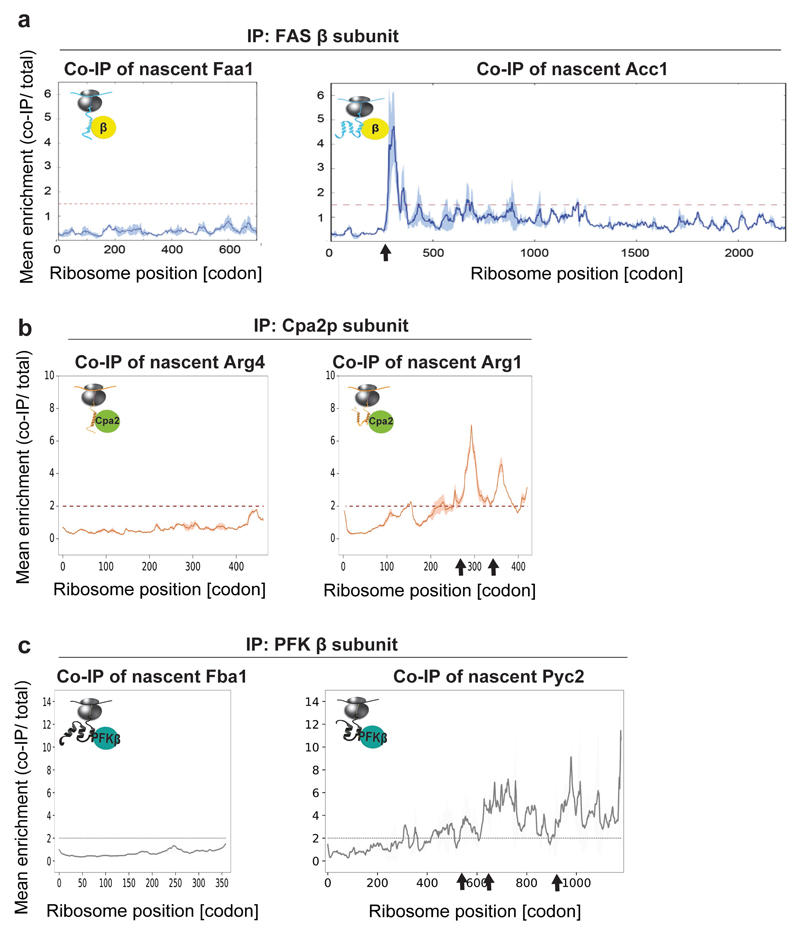
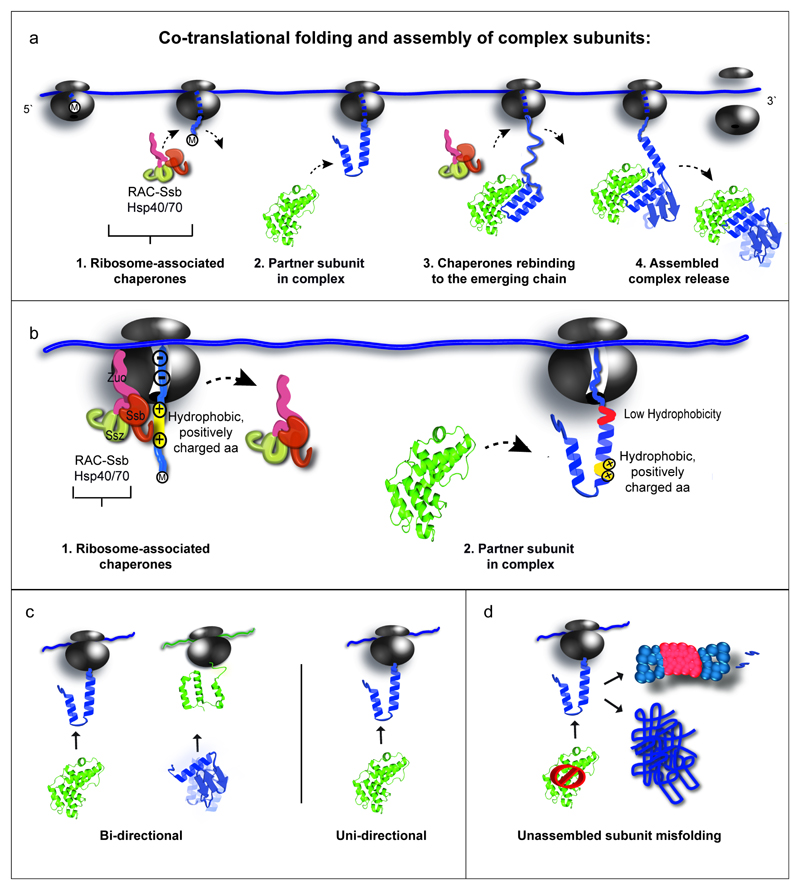
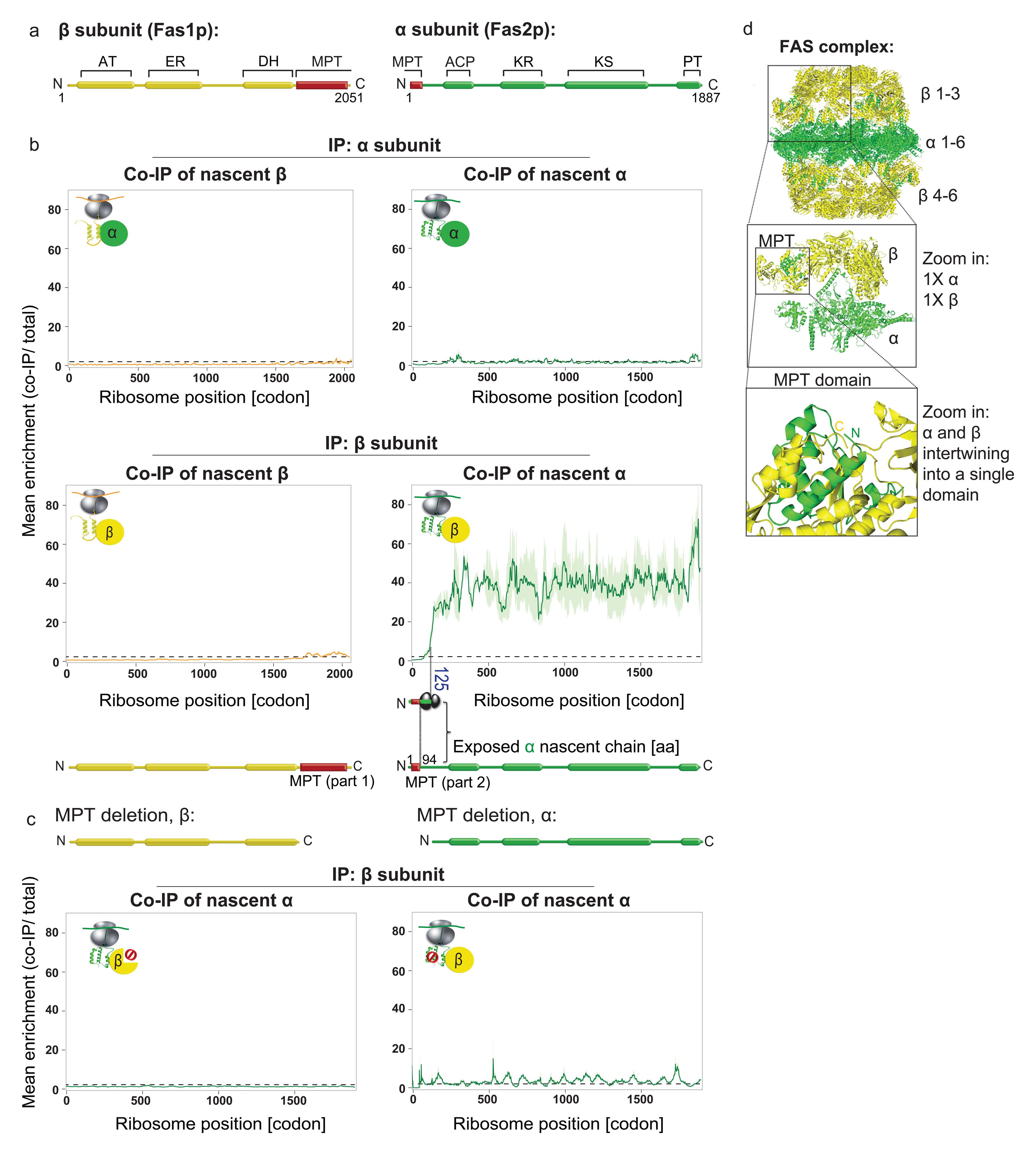
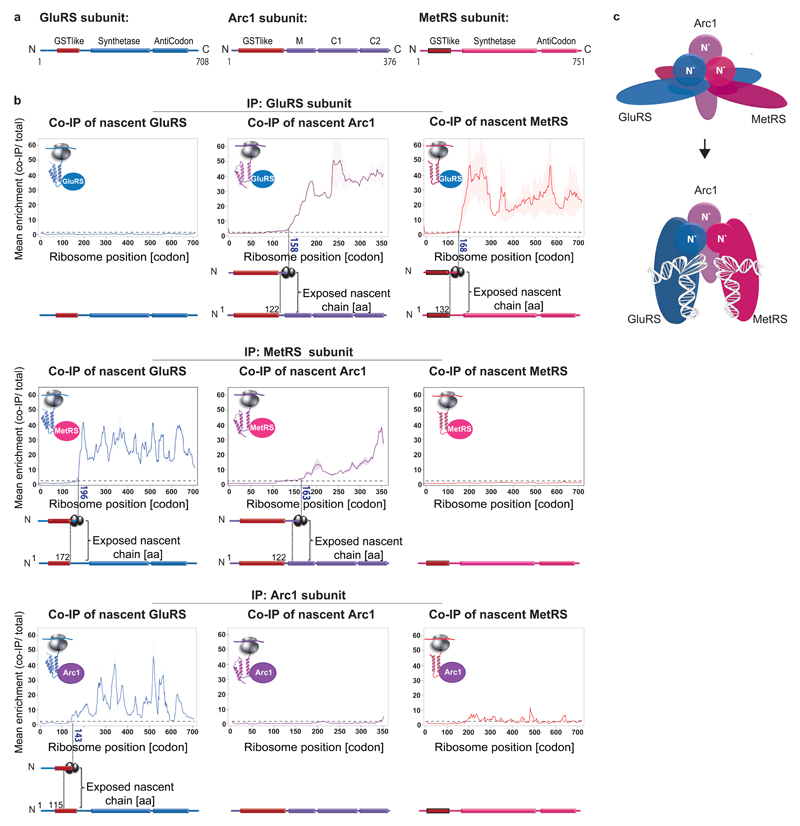
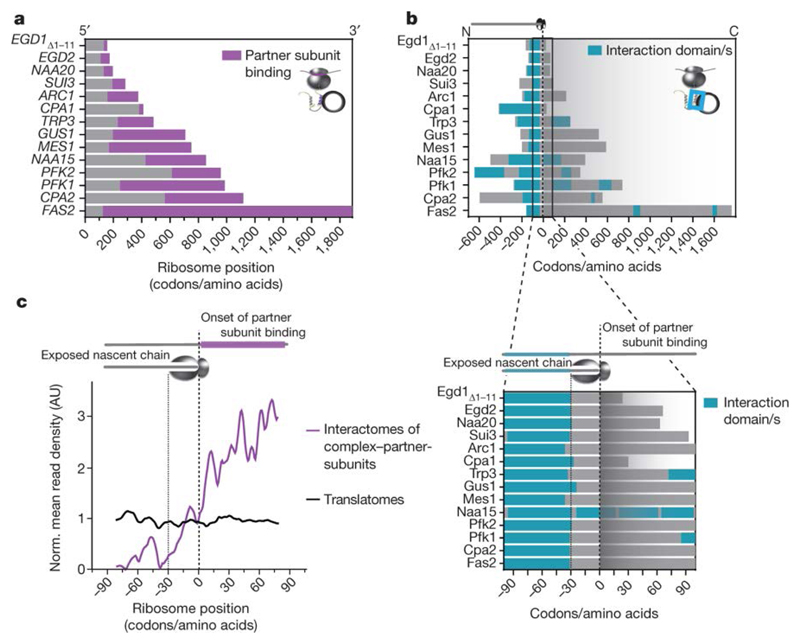
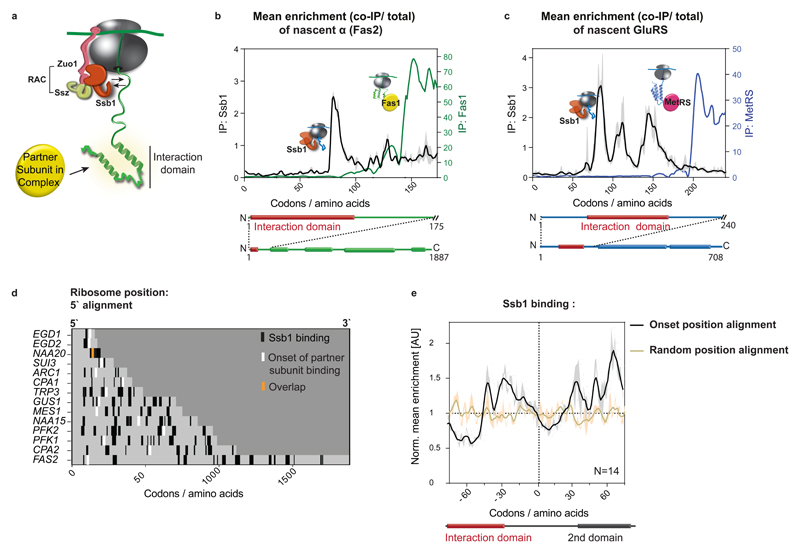
The folding of newly synthesized proteins to the native state is a major challenge within the crowded cellular environment, as non-productive interactions can lead to misfolding, aggregation and degradation1. Cells cope with this challenge by coupling synthesis with polypeptide folding and by using molecular chaperones to safeguard folding cotranslationally2. However, although most of the cellular proteome forms oligomeric assemblies3, little is known about the final step of folding: the assembly of polypeptides into complexes. In prokaryotes, a proof-of-concept study showed that the assembly of heterodimeric luciferase is an organized cotranslational process that is facilitated by spatially confined translation of the subunits encoded on a polycistronic mRNA4. In eukaryotes, however, fundamental differences-such as the rarity of polycistronic mRNAs and different chaperone constellations-raise the question of whether assembly is also coordinated with translation. Here we provide a systematic and mechanistic analysis of the assembly of protein complexes in eukaryotes using ribosome profiling. We determined the in vivo interactions of the nascent subunits from twelve hetero-oligomeric protein complexes of Saccharomyces cerevisiae at near-residue resolution. We find nine complexes assemble cotranslationally; the three complexes that do not show cotranslational interactions are regulated by dedicated assembly chaperones5-7. Cotranslational assembly often occurs uni-directionally, with one fully synthesized subunit engaging its nascent partner subunit, thereby counteracting its propensity for aggregation. The onset of cotranslational subunit association coincides directly with the full exposure of the nascent interaction domain at the ribosomal tunnel exit. The action of the ribosome-associated Hsp70 chaperone Ssb8 is coordinated with assembly. Ssb transiently engages partially synthesized interaction domains and then dissociates before the onset of partner subunit association, presumably to prevent premature assembly interactions. Our study shows that cotranslational subunit association is a prevalent mechanism for the assembly of hetero-oligomers in yeast and indicates that translation, folding and the assembly of protein complexes are integrated processes in eukaryotes.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Protein complexes assemble as they are being made.Nature. 2018 Sep;561(7722):186-187. doi: 10.1038/d41586-018-05905-4. Nature. 2018. PMID: 30194367 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
