

Chemical Tools for Selective Activity Profiling of Endogenously Expressed MMP-14 in Multicellular Models
- PMID: 30160940
- PMCID: PMC6763207
- DOI: 10.1021/acschembio.8b00562
Chemical Tools for Selective Activity Profiling of Endogenously Expressed MMP-14 in Multicellular Models
Abstract
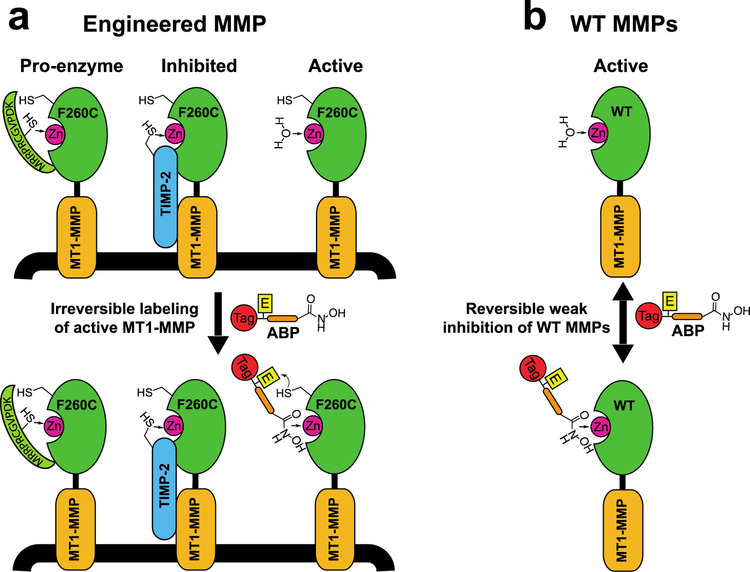
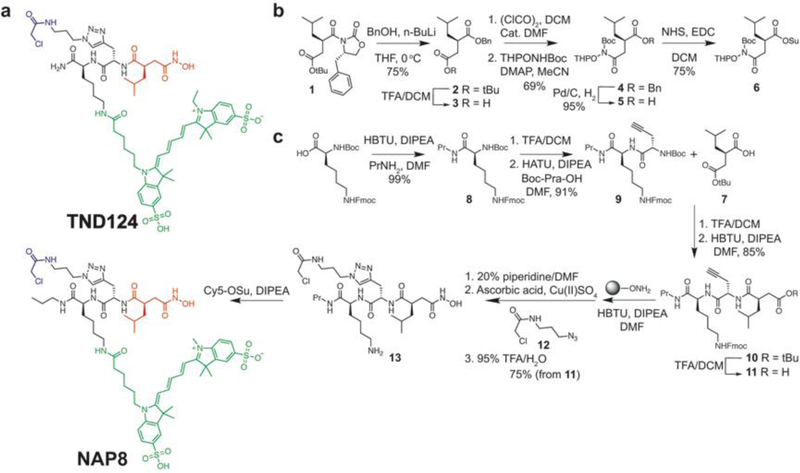
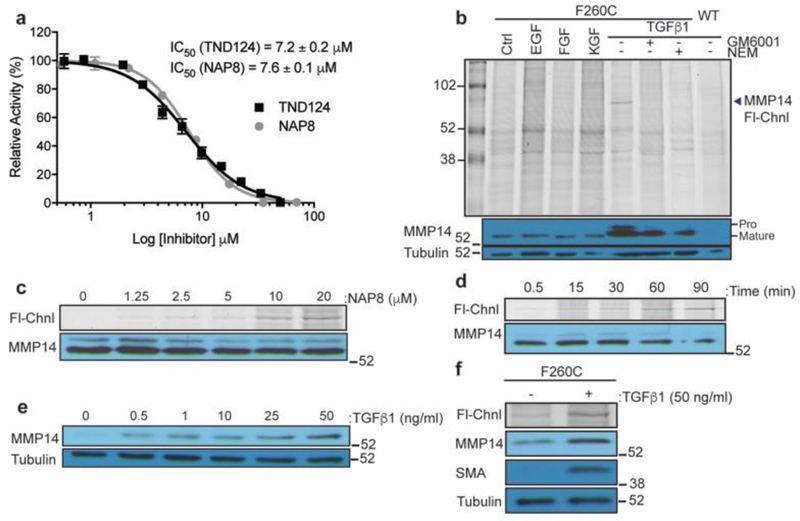
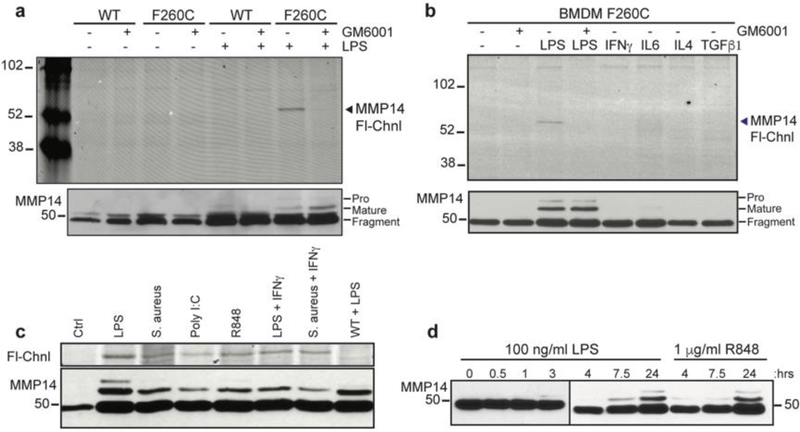
Matrix metalloproteases (MMPs) are a large family of zinc-dependent endopeptidases involved in a diverse set of physiological and pathological processes, most notably in cancer. Current methods for imaging and quantifying MMP activity lack sufficient selectivity and spatiotemporal resolution to allow studies of specific MMP function in vivo. Previously, we reported a strategy for selective targeting of MMPs by engineering a functionally silent cysteine mutation that enables highly specific covalent modification by a designed activity-based probe. Here, we describe the translation of that technology into a mouse model of breast cancer and subsequent demonstration of the utility of the approach for studies of MMP-14 activation in the tumor microenvironment. Using this approach, we find that MMP-14 is active in late stage tumors and is predominantly associated with stromal cell populations that have been activated by specific signaling molecules (e.g., TGFβ) produced by tumor cells. Our data demonstrate the applicability of this approach for studies of MMP function in whole organisms and identify important regulatory mechanisms for MMP-14 activity in the tumor microenvironment.
Figures
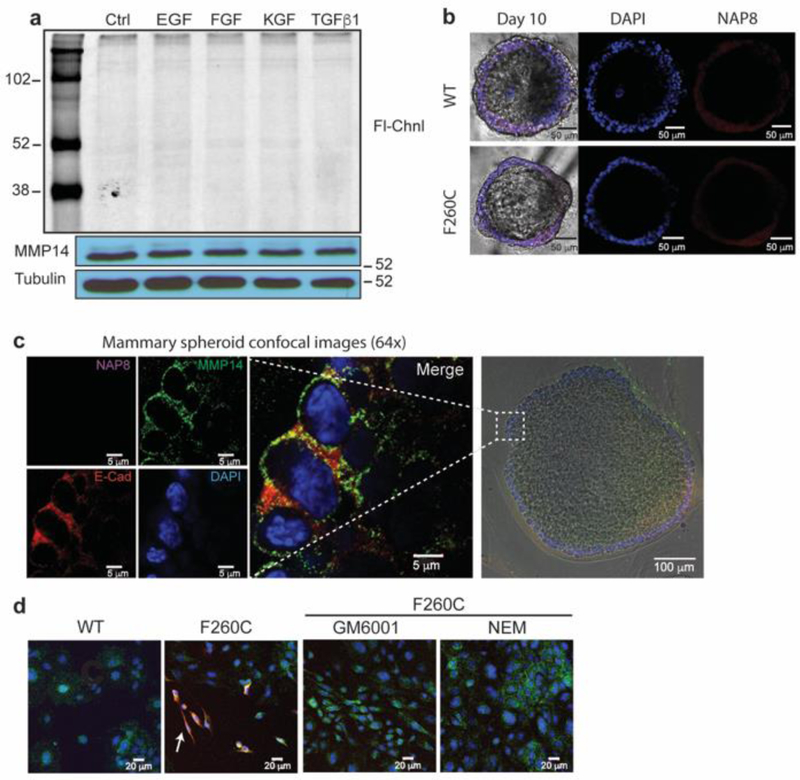
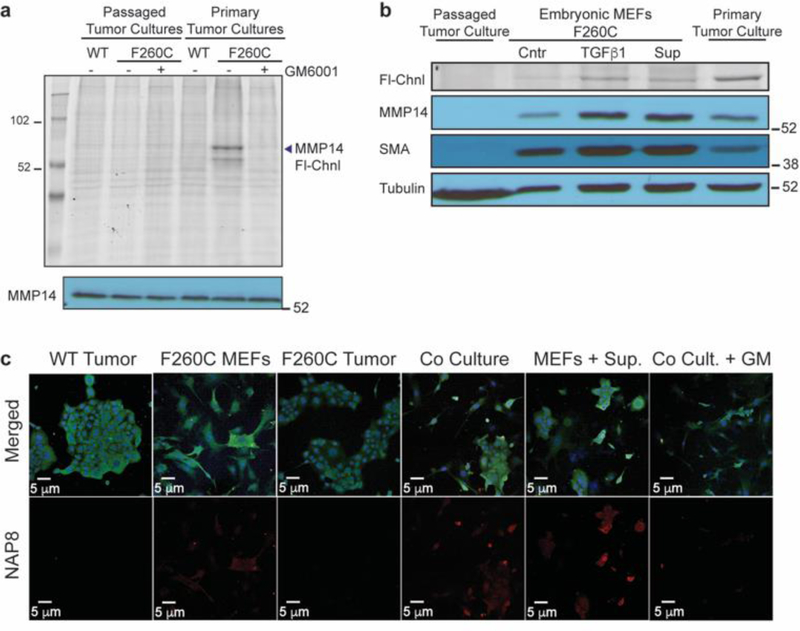
References
-
- Nagase H., Visse R., and Murphy G. (2006) Structure and function of matrix metalloproteinases and TIMPs, Cardiovasc Res 69, 562–573. - PubMed
-
- Rowe RG., and Weiss SJ. (2009) Navigating ECM barriers at the invasive front: the cancer cell-stroma interface, Annu Rev Cell Dev Biol 25, 567–595. - PubMed
-
- Overall CM., and Kleifeld O. (2006) Tumour microenvironment - opinion: validating matrix metalloproteinases as drug targets and anti-targets for cancer therapy, Nat Rev Cancer 6, 227–239. - PubMed
-
- Parks WC., Wilson CL., and Lopez-Boado YS. (2004) Matrix metalloproteinases as modulators of inflammation and innate immunity, Nat Rev Immunol 4, 617–629. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
