

Integrating coalescent species delimitation with analysis of host specificity reveals extensive cryptic diversity despite minimal mitochondrial divergence in the malaria parasite genus Leucocytozoon
- PMID: 30165810
- PMCID: PMC6117968
- DOI: 10.1186/s12862-018-1242-x
Integrating coalescent species delimitation with analysis of host specificity reveals extensive cryptic diversity despite minimal mitochondrial divergence in the malaria parasite genus Leucocytozoon
Abstract
Background: Coalescent methods that use multi-locus sequence data are powerful tools for identifying putatively reproductively isolated lineages, though this approach has rarely been used for the study of microbial groups that are likely to harbor many unrecognized species. Among microbial symbionts, integrating genetic species delimitation methods with trait data that could indicate reproductive isolation, such as host specificity data, has rarely been used despite its potential to inform species limits. Here we test the ability of an integrative approach combining genetic and host specificity data to delimit species within the avian malaria parasite genus Leucocytozoon in central Alaska.
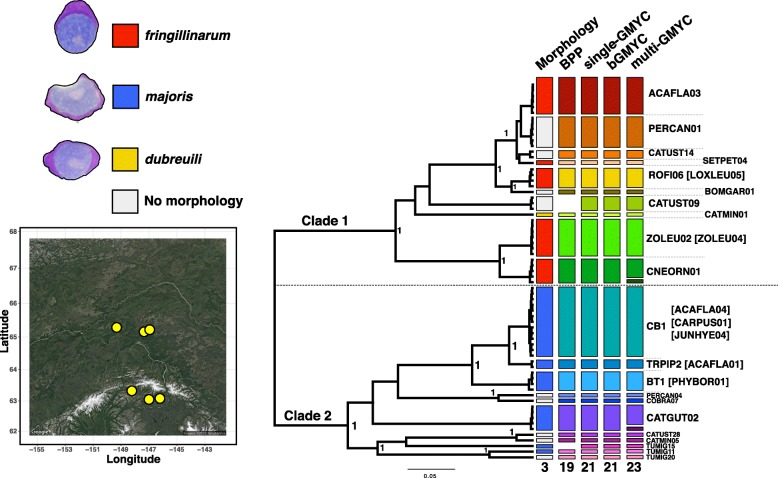
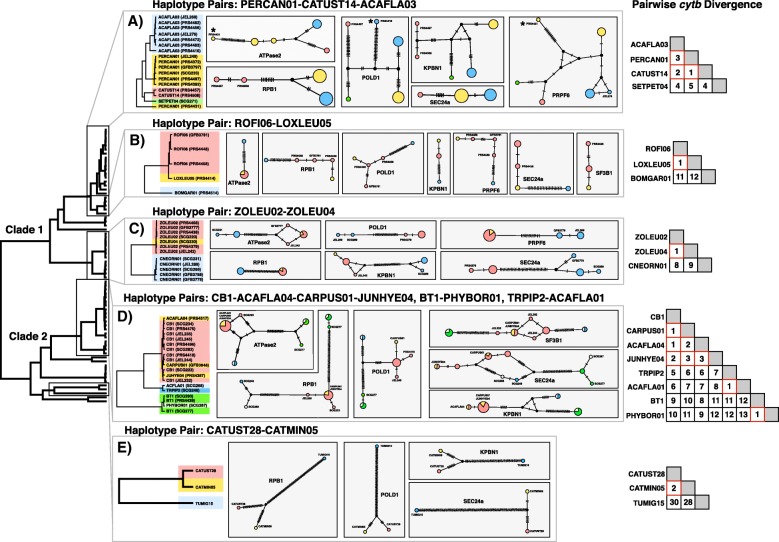
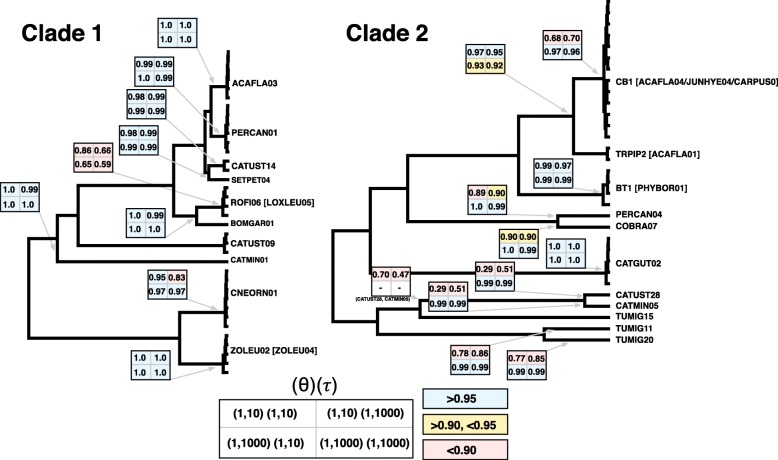
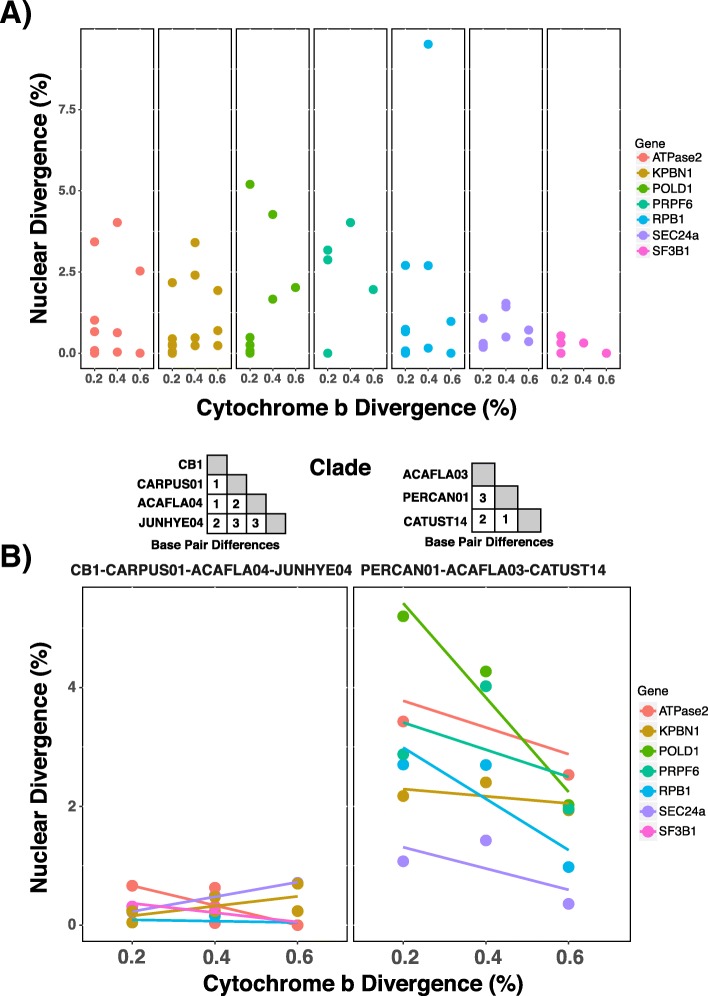
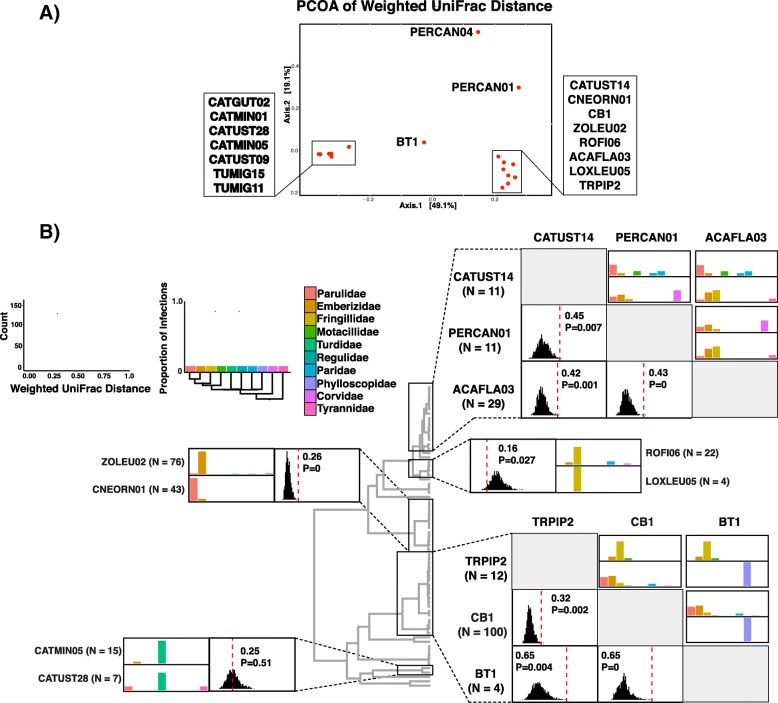
Results: We sequenced seven nuclear loci for 69 Leucocytozoon samples and used multiple species delimitation methods (GMYC and BPP models), tested for differences in host infection patterns among putative species based on 406 individual infections, and characterized parasite morphology. We found that cryptic morphology has masked a highly diverse Leucocytozoon assemblage, with most species delimitation methods recovering support for at least 21 separate species that occur sympatrically and have divergent host infection patterns. Reproductive isolation among putative species appears to have evolved despite low mtDNA divergence, and in one instance two Leucocytozoon cytb haplotypes that differed by a single base pair (~ 0.2% divergence) were supported as separate species. However, there was no consistent association between mtDNA divergence and species limits. Among cytb haplotypes that differed by one to three base pairs we observed idiosyncratic patterns of nuclear and ecological divergence, with cytb haplotype pairs found to be either conspecific, reproductively isolated with no divergence in host specificity, or reproductively isolated with divergent patterns of host specialization.
Conclusion: Integrating multi-locus genetic species delimitation methods and non-traditional ecological data types such as host specificity provide a novel view of the diversity of avian malaria parasites that has been missed previously using morphology and mtDNA barcodes. Species delimitation methods show that Leucocytozoon is highly species-rich in Alaska, and the genus is likely to harbor extraordinary species-level diversity worldwide. Integrating genetic and ecological data will be an important approach for understanding the diversity and evolutionary history of microbial symbionts moving forward.
Keywords: Avian malaria; BPP; Cryptic species; GMYC; Haemosporida; UniFrac.
Conflict of interest statement
Ethics approval
Birds were collected under State of Alaska Department of Fish and Game Scientific Permits 16–013 and 17–092 and approved by the Institutional Animal Care and Use Committee of the American Museum of Natural History.
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Genetic diversity and host specificity varies across three genera of blood parasites in ducks of the Pacific Americas Flyway.PLoS One. 2015 Feb 24;10(2):e0116661. doi: 10.1371/journal.pone.0116661. eCollection 2015. PLoS One. 2015. PMID: 25710468 Free PMC article.
-
Molecular analyses on host-seeking black flies (Diptera: Simuliidae) reveal a diverse assemblage of Leucocytozoon (Apicomplexa: Haemospororida) parasites in an alpine ecosystem.Parasit Vectors. 2015 Jun 25;8:343. doi: 10.1186/s13071-015-0952-9. Parasit Vectors. 2015. PMID: 26108211 Free PMC article.
-
Linkage between nuclear and mitochondrial DNA sequences in avian malaria parasites: multiple cases of cryptic speciation?Evolution. 2004 Jul;58(7):1617-21. doi: 10.1111/j.0014-3820.2004.tb01742.x. Evolution. 2004. PMID: 15341164
-
Within-host speciation of malaria parasites.PLoS One. 2007 Feb 21;2(2):e235. doi: 10.1371/journal.pone.0000235. PLoS One. 2007. PMID: 17311104 Free PMC article.
-
Species limits in avian malaria parasites (Haemosporida): how to move forward in the molecular era.Parasitology. 2014 Sep;141(10):1223-32. doi: 10.1017/S0031182014000560. Epub 2014 May 9. Parasitology. 2014. PMID: 24813385 Review.
Cited by
-
Leucocytozoon infections in tits (Aves, Paridae): blood and tissue stages investigated using an integrative approach.Parasite. 2025;32:13. doi: 10.1051/parasite/2025007. Epub 2025 Feb 21. Parasite. 2025. PMID: 39982000 Free PMC article.
-
Molecular detection and genetic diversity of Leucocytozoon sabrazesi in chickens in Thailand.Sci Rep. 2021 Aug 17;11(1):16686. doi: 10.1038/s41598-021-96241-7. Sci Rep. 2021. PMID: 34404893 Free PMC article.
-
Phylogenomics from transcriptomic "bycatch" clarify the origins and diversity of avian trypanosomes in North America.PLoS One. 2020 Oct 8;15(10):e0240062. doi: 10.1371/journal.pone.0240062. eCollection 2020. PLoS One. 2020. PMID: 33031471 Free PMC article.
-
Population structure of avian malaria parasites.Ecol Evol. 2019 Jun 17;9(13):7741-7751. doi: 10.1002/ece3.5356. eCollection 2019 Jul. Ecol Evol. 2019. PMID: 31346436 Free PMC article.
-
Coinfection rates of avian blood parasites increase with latitude in parapatric host species.Parasitology. 2023 Apr;150(4):329-336. doi: 10.1017/S0031182022001792. Epub 2023 Jan 4. Parasitology. 2023. PMID: 36597832 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
