

Reprogramming of the antimycin NRPS-PKS assembly lines inspired by gene evolution
- PMID: 30166552
- PMCID: PMC6117356
- DOI: 10.1038/s41467-018-05877-z
Reprogramming of the antimycin NRPS-PKS assembly lines inspired by gene evolution
Abstract
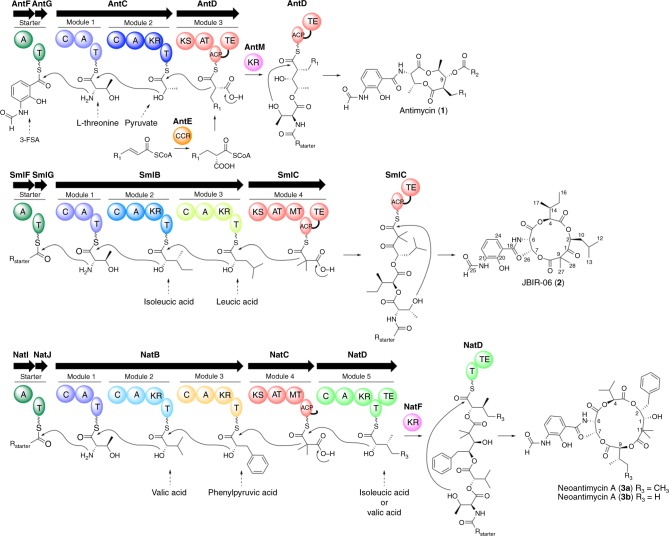
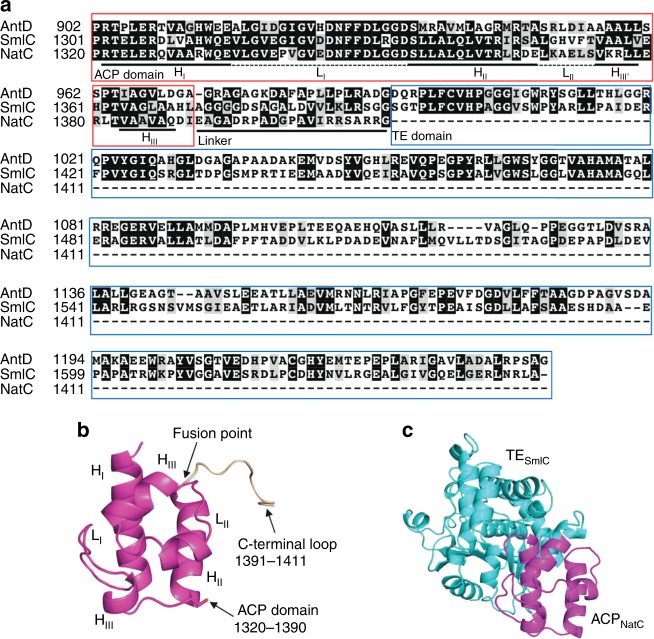
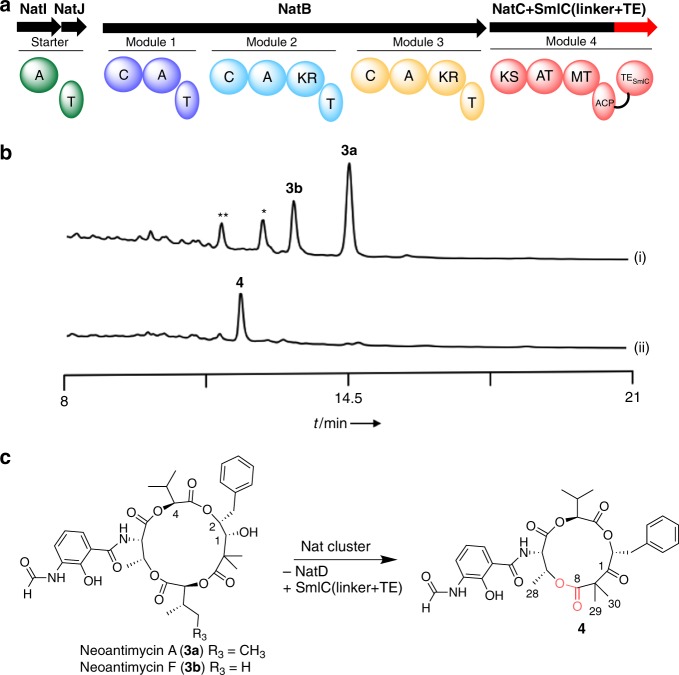
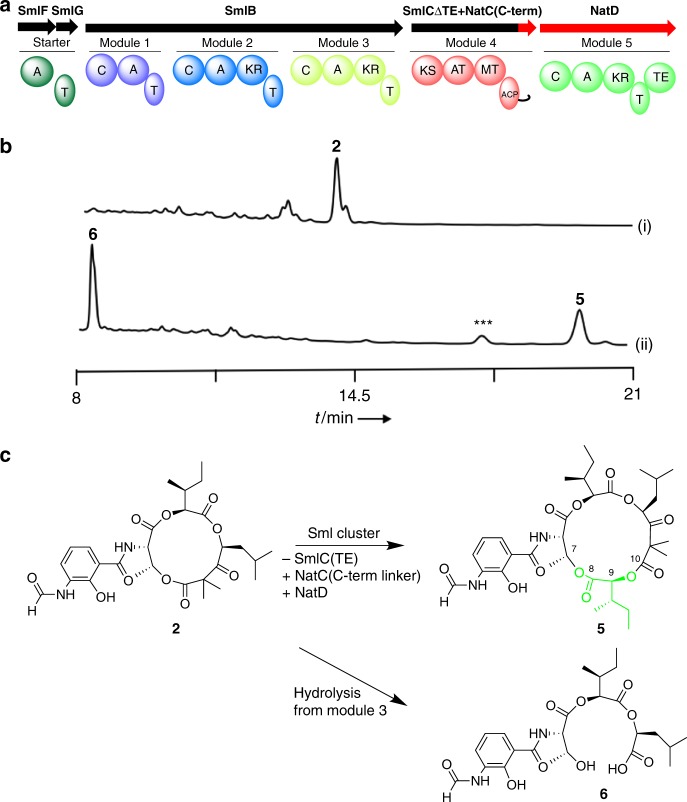
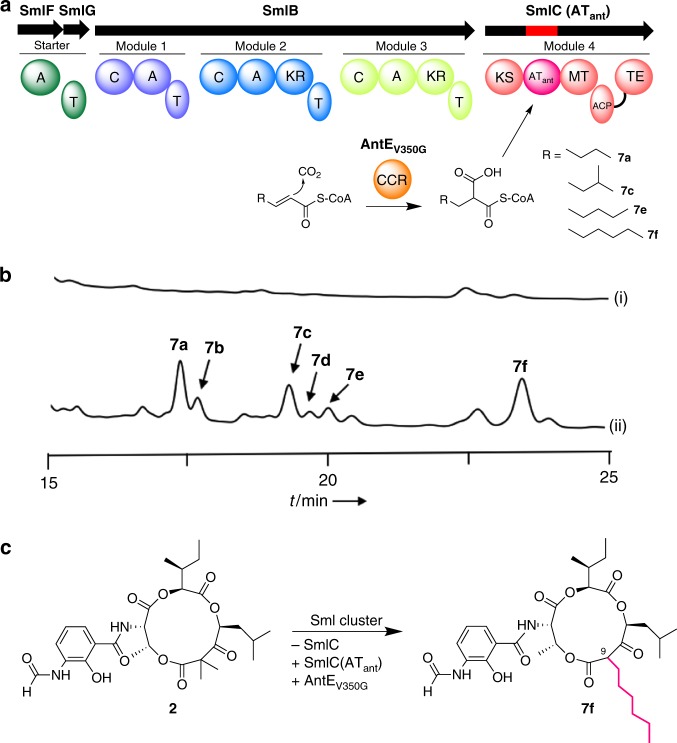
Reprogramming of the NRPS/PKS assembly line is an attractive method for the production of new bioactive molecules. However, it is usually hampered by the loss of intimate domain/module interactions required for the precise control of chain transfer and elongation reactions. In this study, we first establish heterologous expression systems of the unique antimycin-type cyclic depsipeptides: JBIR-06 (tri-lactone) and neoantimycin (tetra-lactone), and engineer their biosyntheses by taking advantage of bioinformatic analyses and evolutionary insights. As a result, we successfully accomplish three manipulations: (i) ring contraction of neoantimycin (from tetra-lactone to tri-lactone), (ii) ring expansion of JBIR-06 (from tri-lactone to tetra-lactone), and (iii) alkyl chain diversification of JBIR-06 by the incorporation of various alkylmalonyl-CoA extender units, to generate a set of unnatural derivatives in practical yields. This study presents a useful strategy for engineering NRPS-PKS module enzymes, based on nature's diversification of the domain and module organizations.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
