

Functional features defining the efficacy of cholesterol-conjugated, self-deliverable, chemically modified siRNAs
- PMID: 30169779
- PMCID: PMC6237813
- DOI: 10.1093/nar/gky745
Functional features defining the efficacy of cholesterol-conjugated, self-deliverable, chemically modified siRNAs
Abstract
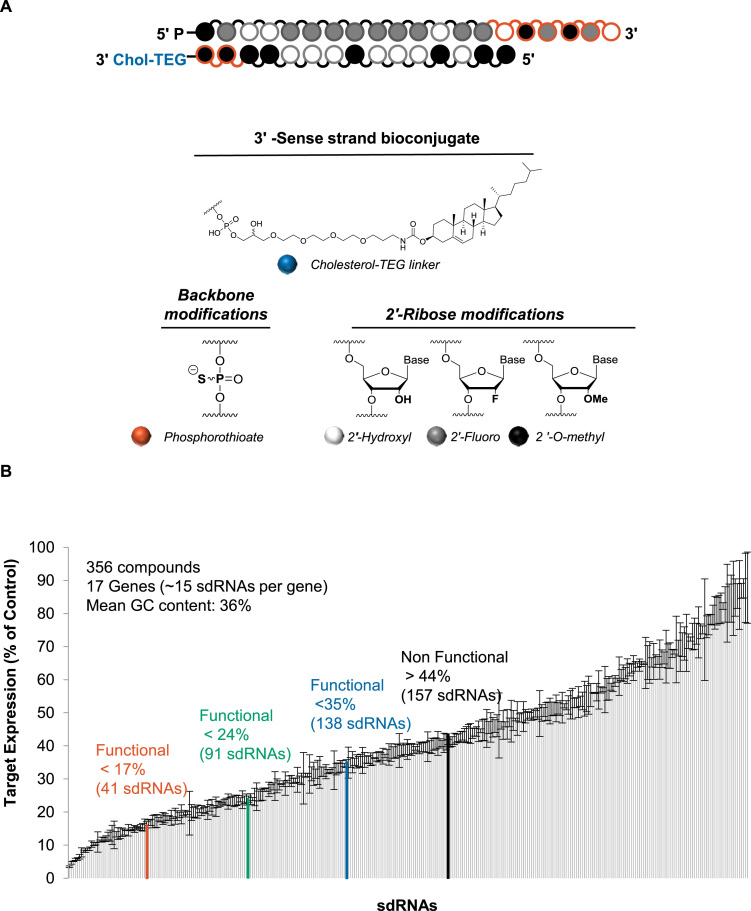
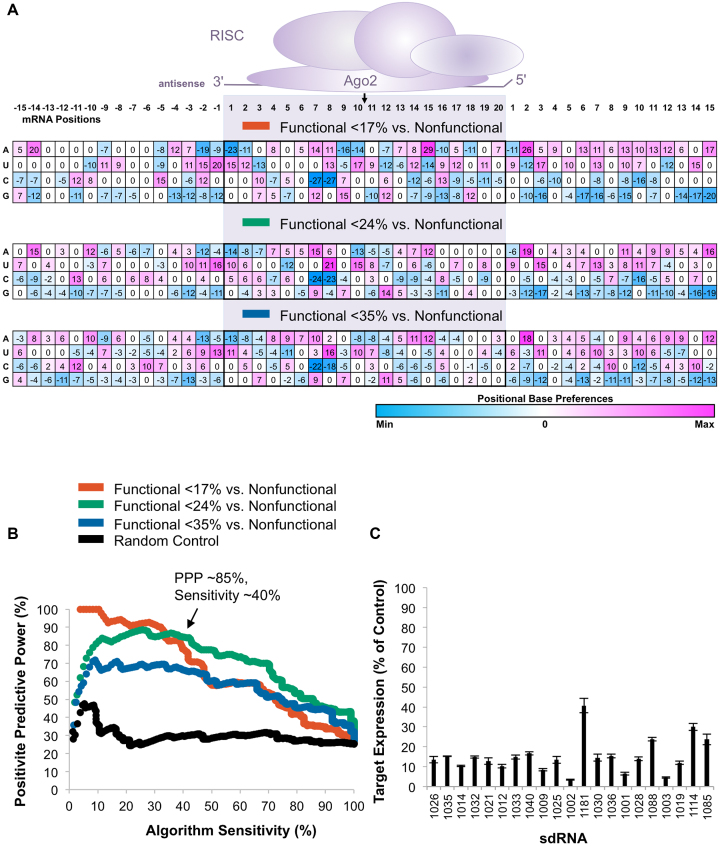
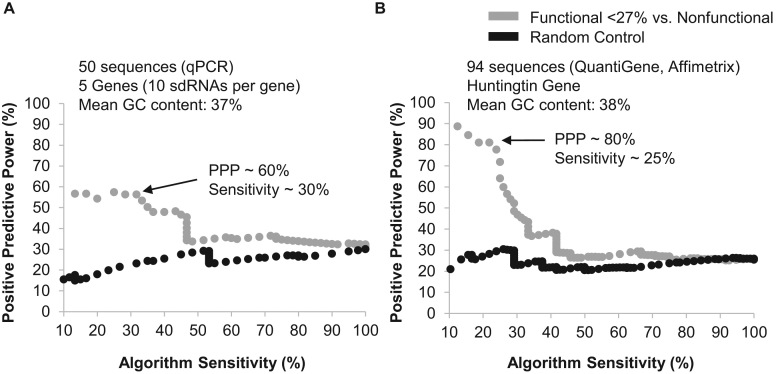
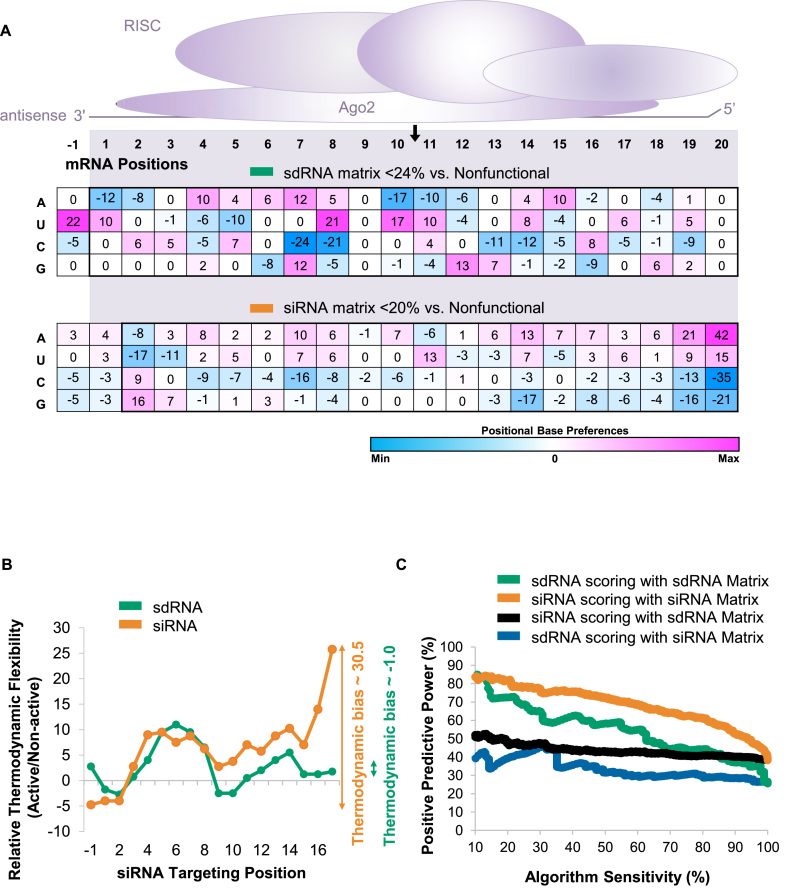
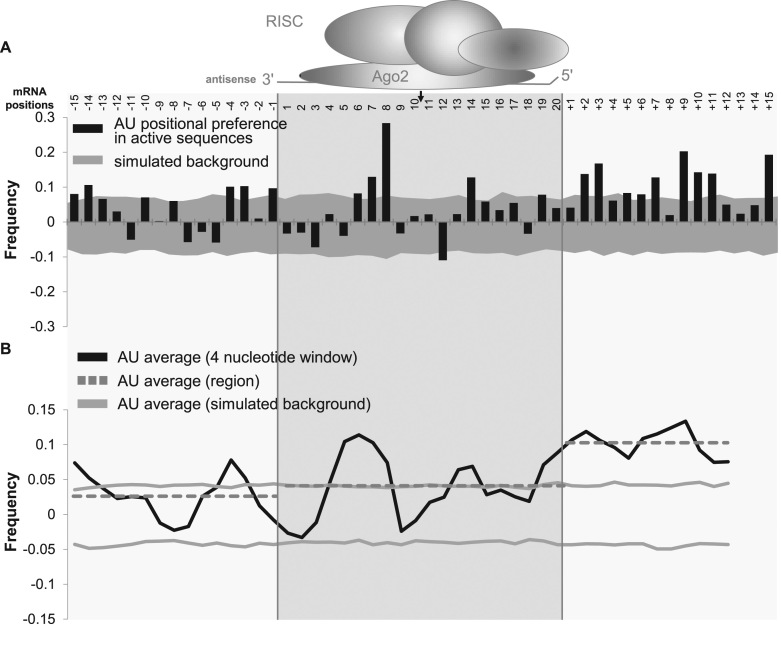
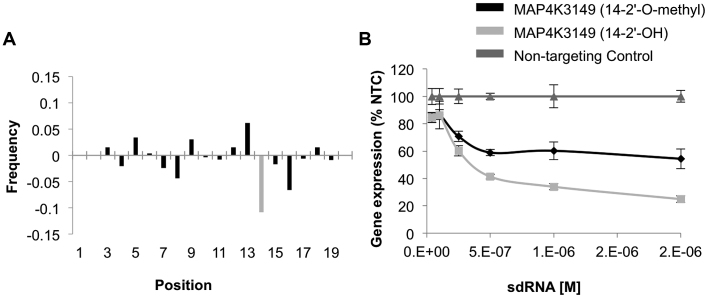
Progress in oligonucleotide chemistry has produced a shift in the nature of siRNA used, from formulated, minimally modified siRNAs, to unformulated, heavily modified siRNA conjugates. The introduction of extensive chemical modifications is essential for conjugate-mediated delivery. Modifications have a significant impact on siRNA efficacy through interference with recognition and processing by RNAi enzymatic machinery, severely restricting the sequence space available for siRNA design. Many algorithms available publicly can successfully predict the activity of non-modified siRNAs, but the efficiency of the algorithms for designing heavily modified siRNAs has never been systematically evaluated experimentally. Here we screened 356 cholesterol-conjugated siRNAs with extensive modifications and developed a linear regression-based algorithm that effectively predicts siRNA activity using two independent datasets. We further demonstrate that predictive determinants for modified and non-modified siRNAs differ substantially. The algorithm developed from the non-modified siRNAs dataset has no predictive power for modified siRNAs and vice versa. In the context of heavily modified siRNAs, the introduction of chemical asymmetry fully eliminates the requirement for thermodynamic bias, the major determinant for non-modified siRNA efficacy. Finally, we demonstrate that in addition to the sequence of the target site, the accessibility of the neighboring 3' region significantly contributes to siRNA efficacy.
Figures
References
-
- Dowdy S.F. Overcoming cellular barriers for RNA therapeutics. Nat. Biotechnol. 2017; 35:222–229. - PubMed
-
- Huesken D., Lange J.A., Mickanin C., Weiler J., Asselbergs F., Warner J., Meloon B., Engel S., Rosenberg A., Cohen D. et al. Design of a genome-wide siRNA library using an artificial neural network. Nat. Biotechnol. 2005; 23:995–1001. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
