

The major nectar protein of Brassica rapa is a non-specific lipid transfer protein, BrLTP2.1, with strong antifungal activity
- PMID: 30169819
- PMCID: PMC6255697
- DOI: 10.1093/jxb/ery319
The major nectar protein of Brassica rapa is a non-specific lipid transfer protein, BrLTP2.1, with strong antifungal activity
Abstract
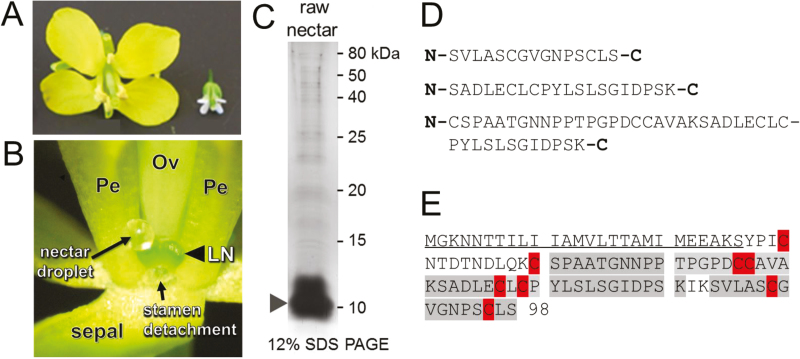
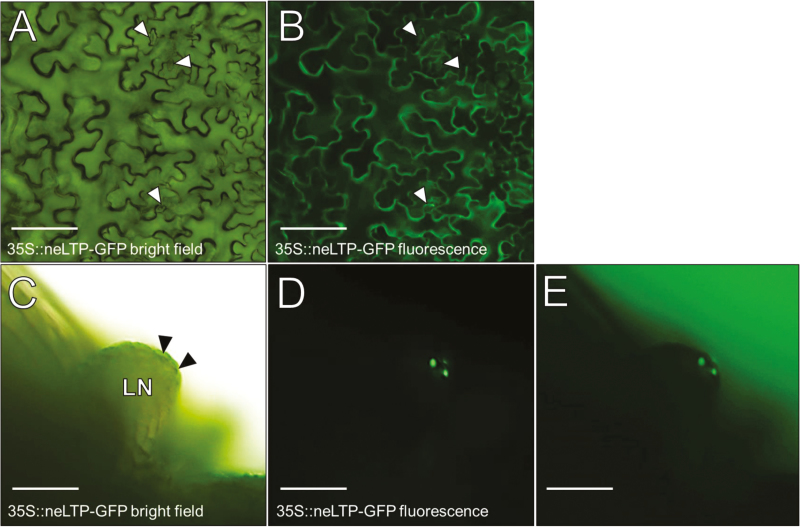
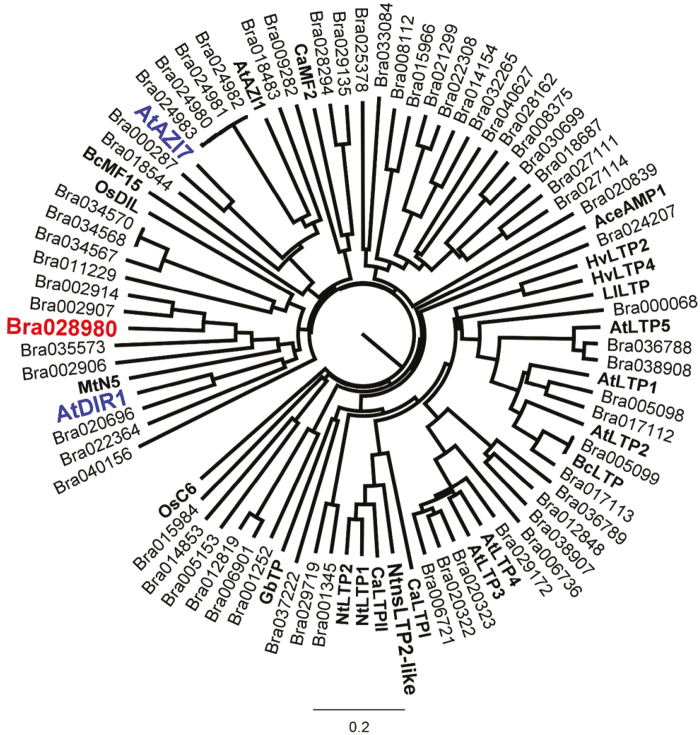
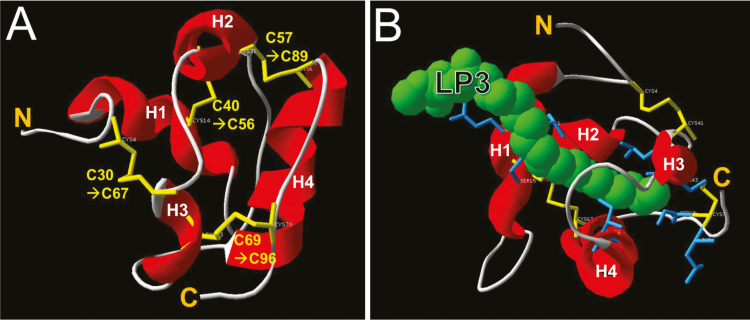
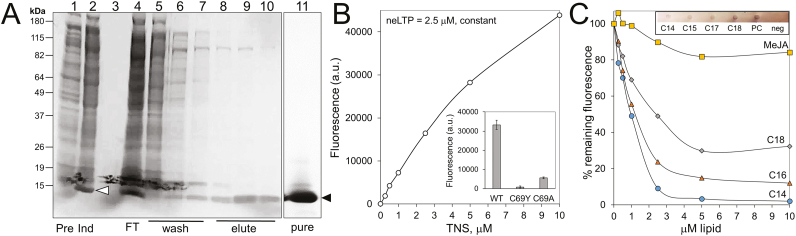
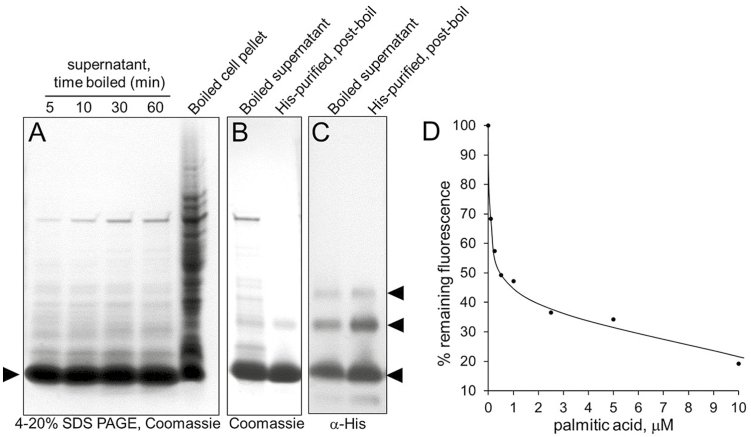
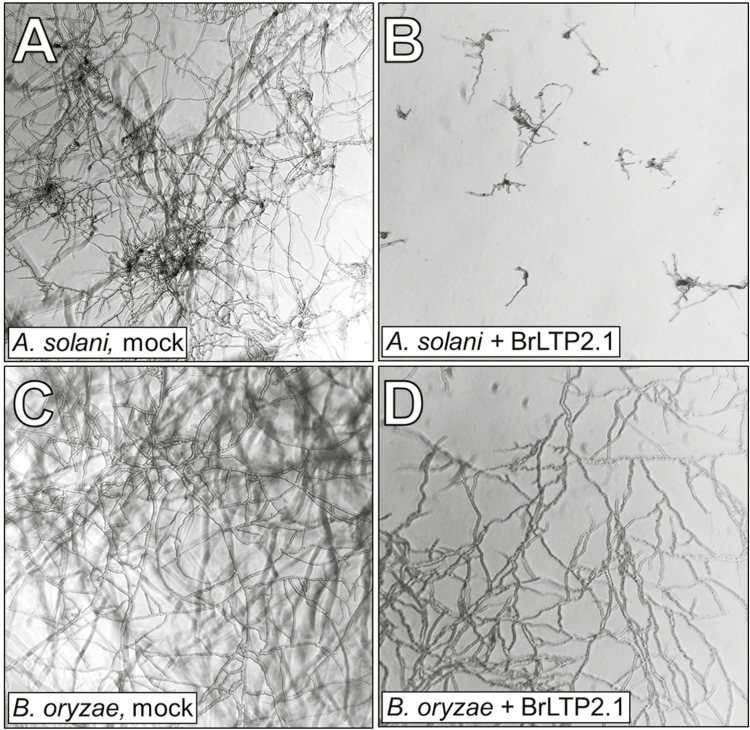
Nectar is one of the key rewards mediating plant-mutualist interactions. In addition to sugars, nectars often contain many other compounds with important biological functions, including proteins. This study was undertaken to assess the proteinaceous content of Brassica rapa nectar. SDS-PAGE analysis of raw B. rapa nectar revealed the presence of ~10 proteins, with a major band at ~10 kDa. This major band was found to contain a non-specific lipid transfer protein encoded by B. rapa locus Bra028980 and subsequently termed BrLTP2.1. Sequence analysis of BrLTP2.1 predicted the presence of a signal peptide required for secretion from the cell, eight cysteines, and a mature molecular mass of 7.3 kDa. Constitutively expressed BrLTP2.1-GFP in Arabidopsis displayed accumulation patterns consistent with secretion from nectary cells. BrLTP2.1 was also found to have relatively high sequence similarity to non-specific lipid-transfer proteins with known functions in plant defense, including Arabidopsis DIR1. Heterologously expressed and purified BrLTP2.1 was extremely heat stable and bound strongly to saturated free fatty acids, but not methyl jasmonate. Recombinant BrLTP2.1 also had direct antimicrobial activity against an extensive range of plant pathogens, being particularly effective against necrotrophic fungi. Taken together, these results suggest that BrLTP2.1 may function to prevent microbial growth in nectars.
Figures
Similar articles
-
A cell wall invertase controls nectar volume and sugar composition.Plant J. 2021 Aug;107(4):1016-1028. doi: 10.1111/tpj.15357. Epub 2021 Jul 29. Plant J. 2021. PMID: 34048120
-
Genome-wide identification, phylogeny, evolution, and expression patterns of MtN3/saliva/SWEET genes and functional analysis of BcNS in Brassica rapa.BMC Genomics. 2018 Mar 2;19(1):174. doi: 10.1186/s12864-018-4554-8. BMC Genomics. 2018. PMID: 29499648 Free PMC article.
-
Three genes encoding AOP2, a protein involved in aliphatic glucosinolate biosynthesis, are differentially expressed in Brassica rapa.J Exp Bot. 2015 Oct;66(20):6205-18. doi: 10.1093/jxb/erv331. Epub 2015 Jul 17. J Exp Bot. 2015. PMID: 26188204 Free PMC article.
-
Genome-wide identification of glucosinolate synthesis genes in Brassica rapa.FEBS J. 2009 Jul;276(13):3559-74. doi: 10.1111/j.1742-4658.2009.07076.x. Epub 2009 May 18. FEBS J. 2009. PMID: 19456863
-
Identification of differential gene expression in Brassica rapa nectaries through expressed sequence tag analysis.PLoS One. 2010 Jan 20;5(1):e8782. doi: 10.1371/journal.pone.0008782. PLoS One. 2010. PMID: 20098697 Free PMC article.
Cited by
-
Lipid transfer protein VAS inhibits the hypersensitive response via reactive oxygen species signaling in Nicotiana benthamiana.J Exp Bot. 2025 Feb 25;76(4):1285-1299. doi: 10.1093/jxb/erae473. J Exp Bot. 2025. PMID: 39921679 Free PMC article.
-
Specialized metabolites present in Camellia reticulata nectar inhibit the growth of nectar-inhabiting microorganisms.Front Plant Sci. 2025 Mar 4;16:1557228. doi: 10.3389/fpls.2025.1557228. eCollection 2025. Front Plant Sci. 2025. PMID: 40104037 Free PMC article.
-
Lipid transfer protein StLTPa enhances potato disease resistance against different pathogens by binding and disturbing the integrity of pathogens plasma membrane.Plant Biotechnol J. 2024 Jul;22(7):1913-1925. doi: 10.1111/pbi.14310. Epub 2024 Feb 16. Plant Biotechnol J. 2024. PMID: 38366362 Free PMC article.
-
Nectar compounds impact bacterial and fungal growth and shift community dynamics in a nectar analog.Environ Microbiol Rep. 2023 Jun;15(3):170-180. doi: 10.1111/1758-2229.13139. Epub 2023 Feb 13. Environ Microbiol Rep. 2023. PMID: 36779256 Free PMC article.
-
Structural and Functional Characterization of New Lipid Transfer Proteins with Chitin-Binding Properties: Insights from Protein Structure Prediction, Molecular Docking, and Antifungal Activity.Biochemistry. 2024 Jul 16;63(14):1824-1836. doi: 10.1021/acs.biochem.4c00124. Epub 2024 Jul 5. Biochemistry. 2024. PMID: 38968244 Free PMC article.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment search tool. Journal of Molecular Biology 215, 403–410. - PubMed
-
- Bakan B, Hamberg M, Perrocheau L, Maume D, Rogniaux H, Tranquet O, Rondeau C, Blein JP, Ponchet M, Marion D. 2006. Specific adduction of plant lipid transfer protein by an allene oxide generated by 9-lipoxygenase and allene oxide synthase. The Journal of Biological Chemistry 281, 38981–38988. - PubMed
-
- Baker HG. 1978. Chemical aspects of the pollination of woody plants in the tropics. In: Tomlinson PB, Zimmerman M, eds. Tropical trees as living systems. New York: Cambridge University Press, 57–82.
-
- Bender RL, Fekete ML, Klinkenberg PM, et al. . 2013. PIN6 is required for nectary auxin response and short stamen development. The Plant Journal 74, 893–904. - PubMed
-
- Bender R, Klinkenberg P, Jiang Z, Bauer B, Karypis G, Nguyen N, Perera MADN, Nikolau BJ, Carter CJ. 2012. Functional genomics of nectar production in the Brassicaceae. Flora 207, 491–496.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
