

Lung γδ T Cells Mediate Protective Responses during Neonatal Influenza Infection that Are Associated with Type 2 Immunity
- PMID: 30170813
- PMCID: PMC6345262
- DOI: 10.1016/j.immuni.2018.07.011
Lung γδ T Cells Mediate Protective Responses during Neonatal Influenza Infection that Are Associated with Type 2 Immunity
Abstract
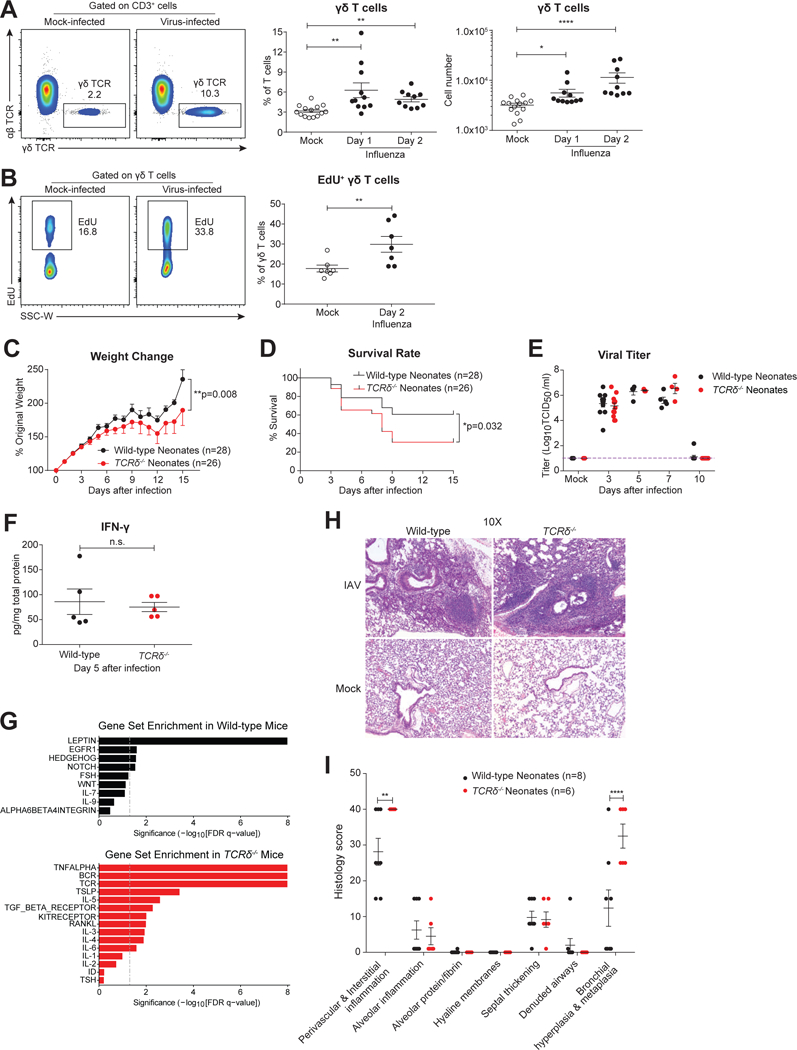
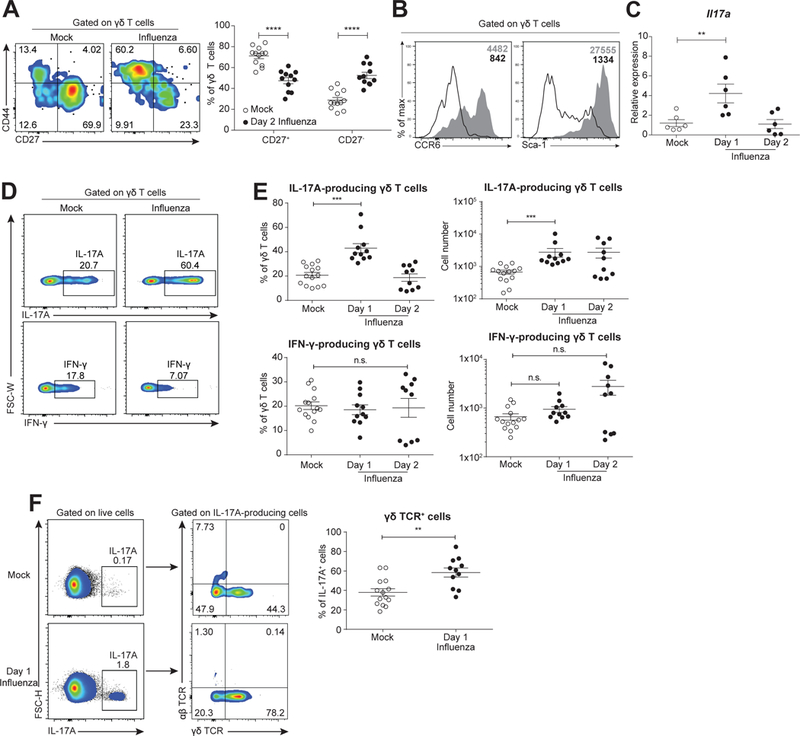
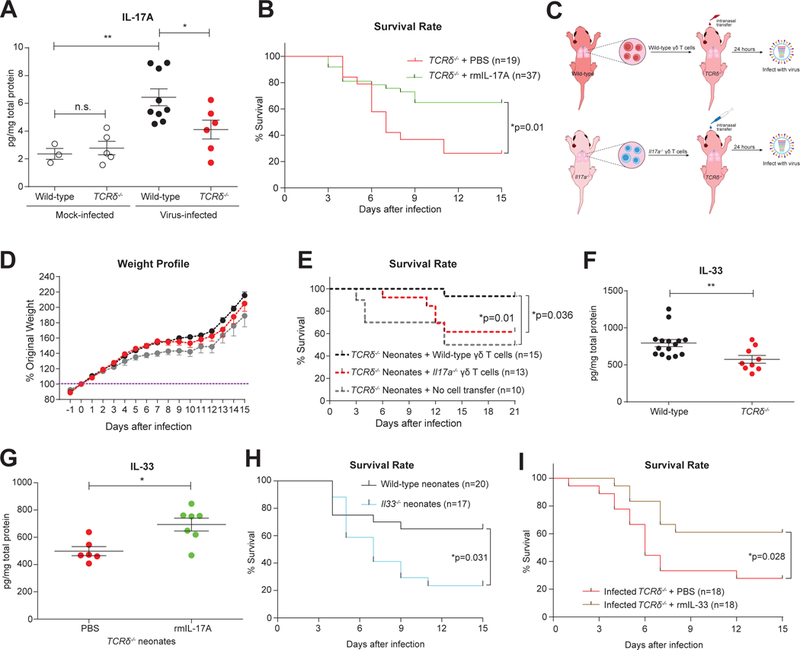
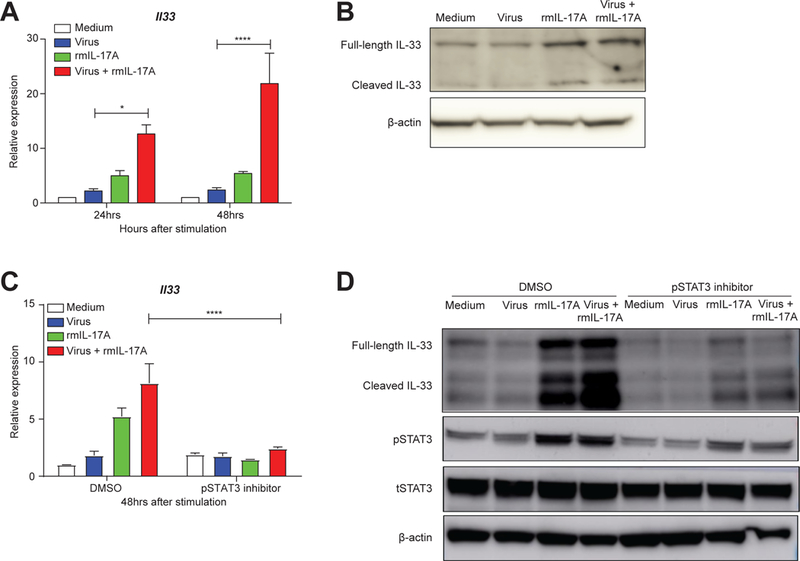
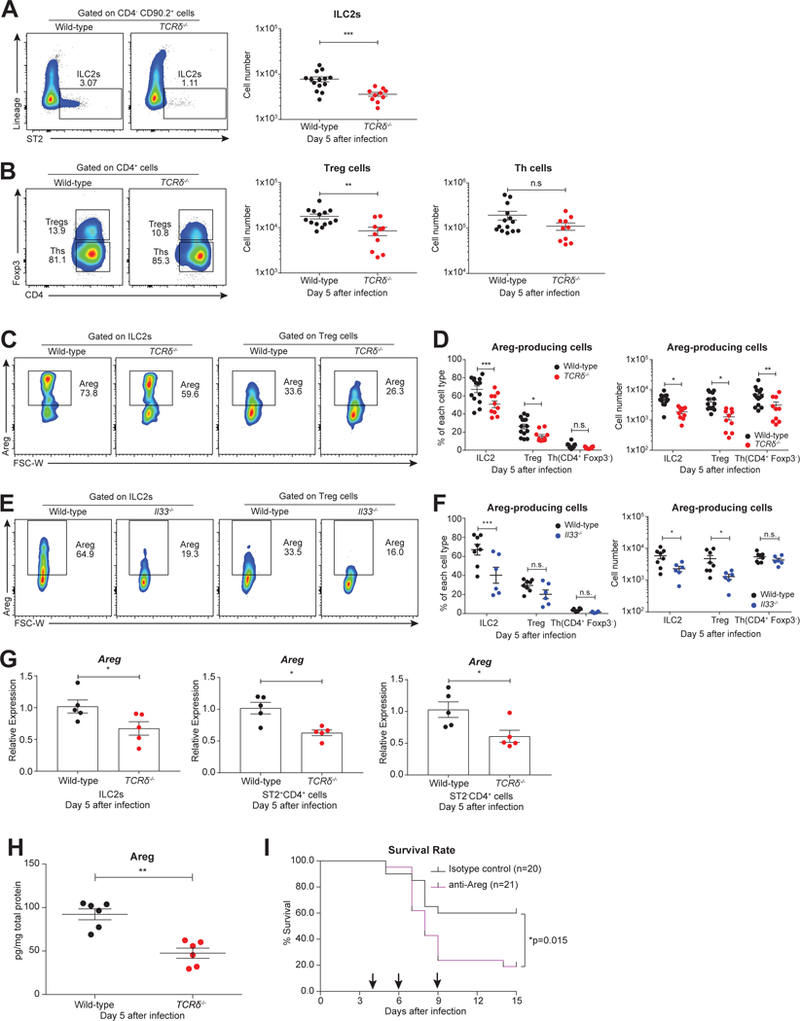
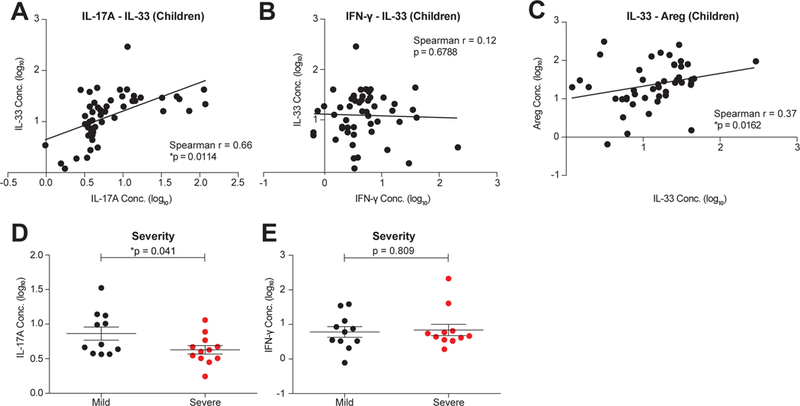
Compared to adults, infants suffer higher rates of hospitalization, severe clinical complications, and mortality due to influenza infection. We found that γδ T cells protected neonatal mice against mortality during influenza infection. γδ T cell deficiency did not alter viral clearance or interferon-γ production. Instead, neonatal influenza infection induced the accumulation of interleukin-17A (IL-17A)-producing γδ T cells, which was associated with IL-33 production by lung epithelial cells. Neonates lacking IL-17A-expressing γδ T cells or Il33 had higher mortality upon influenza infection. γδ T cells and IL-33 promoted lung infiltration of group 2 innate lymphoid cells and regulatory T cells, resulting in increased amphiregulin secretion and tissue repair. In influenza-infected children, IL-17A, IL-33, and amphiregulin expression were correlated, and increased IL-17A levels in nasal aspirates were associated with better clinical outcomes. Our results indicate that γδ T cells are required in influenza-infected neonates to initiate protective immunity and mediate lung homeostasis.
Keywords: IL-17A; IL-33; amphiregulin; children; neonatal influenza infection; type 2 immunity; γδ T cells.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Host-derived lipids orchestrate pulmonary γδ T cell response to provide early protection against influenza virus infection.Nat Commun. 2021 Mar 26;12(1):1914. doi: 10.1038/s41467-021-22242-9. Nat Commun. 2021. PMID: 33772013 Free PMC article.
-
Protective role of γδ T cells in cigarette smoke and influenza infection.Mucosal Immunol. 2018 May;11(3):894-908. doi: 10.1038/mi.2017.93. Epub 2017 Nov 1. Mucosal Immunol. 2018. PMID: 29091081 Free PMC article.
-
Neuronal-Activated ILC2s Promote IL-17A Production in Lung γδ T Cells During Sepsis.Front Immunol. 2021 Apr 30;12:670676. doi: 10.3389/fimmu.2021.670676. eCollection 2021. Front Immunol. 2021. PMID: 33995408 Free PMC article.
-
Role of γδ T cells in controlling viral infections with a focus on influenza virus: implications for designing novel therapeutic approaches.Virol J. 2020 Nov 12;17(1):174. doi: 10.1186/s12985-020-01449-0. Virol J. 2020. PMID: 33183352 Free PMC article. Review.
-
Regulation and function of IL-17A- and IL-22-producing γδ T cells.Cell Mol Life Sci. 2011 Jul;68(14):2371-90. doi: 10.1007/s00018-011-0700-z. Epub 2011 May 15. Cell Mol Life Sci. 2011. PMID: 21573786 Free PMC article. Review.
Cited by
-
A Distinctive γδ T Cell Repertoire in NOD Mice Weakens Immune Regulation and Favors Diabetic Disease.Biomolecules. 2022 Oct 1;12(10):1406. doi: 10.3390/biom12101406. Biomolecules. 2022. PMID: 36291615 Free PMC article.
-
COVID-19: Proposing a Ketone-Based Metabolic Therapy as a Treatment to Blunt the Cytokine Storm.Oxid Med Cell Longev. 2020 Sep 9;2020:6401341. doi: 10.1155/2020/6401341. eCollection 2020. Oxid Med Cell Longev. 2020. PMID: 33014275 Free PMC article. Review.
-
Ketogenic diet activates protective γδ T cell responses against influenza virus infection.Sci Immunol. 2019 Nov 15;4(41):eaav2026. doi: 10.1126/sciimmunol.aav2026. Sci Immunol. 2019. PMID: 31732517 Free PMC article.
-
Hematopoiesis: A Layered Organization Across Chordate Species.Front Cell Dev Biol. 2020 Dec 16;8:606642. doi: 10.3389/fcell.2020.606642. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33392196 Free PMC article. Review.
-
Streptococcus agalactiae and Escherichia coli Induce Distinct Effector γδ T Cell Responses During Neonatal Sepsis.bioRxiv [Preprint]. 2024 Mar 16:2023.10.02.560561. doi: 10.1101/2023.10.02.560561. bioRxiv. 2024. Update in: iScience. 2024 Apr 05;27(5):109669. doi: 10.1016/j.isci.2024.109669. PMID: 37873122 Free PMC article. Updated. Preprint.
References
-
- Adkins B, Leclerc C, and Marshall-Clarke S (2004). Neonatal adaptive immunity comes of age. Nat. Rev. Immunol. 4, 553–564. - PubMed
-
- Bates D, Mächler M, Bolker B, and Walker S (2015). Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software, Articles 67, 1–48.
-
- Bhat N, Wright JG, Broder KR, Murray EL, Greenberg ME, Glover MJ, Likos AM, Posey DL, Klimov A, Lindstrom SE, et al. (2005). Influenza-Associated Deaths among Children in the United States, 2003–2004. N. Engl. J. Med. 353, 2559–2567. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
