

Differential biochemical properties of three canonical Dps proteins from the cyanobacterium Nostoc punctiforme suggest distinct cellular functions
- PMID: 30171072
- PMCID: PMC6204913
- DOI: 10.1074/jbc.RA118.002425
Differential biochemical properties of three canonical Dps proteins from the cyanobacterium Nostoc punctiforme suggest distinct cellular functions
Abstract
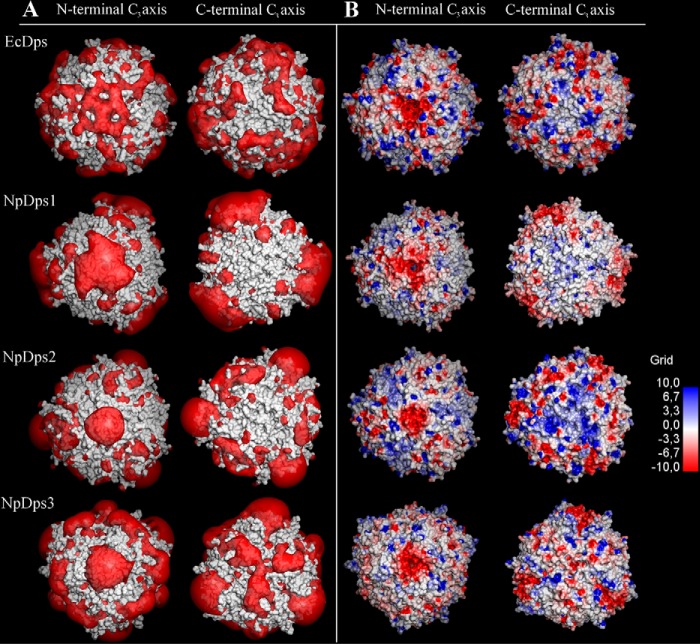
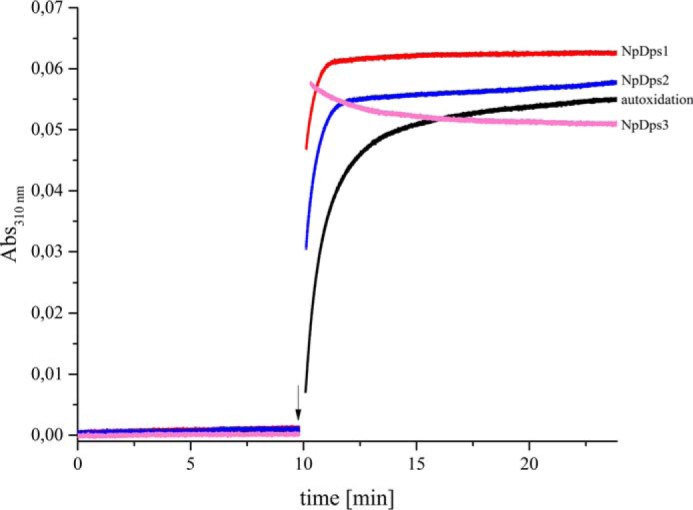
DNA-binding proteins from starved cells (Dps, EC: 1.16.3.1) have a variety of different biochemical activities such as DNA-binding, iron sequestration, and H2O2 detoxification. Most bacteria commonly feature one or two Dps enzymes, whereas the cyanobacterium Nostoc punctiforme displays an unusually high number of five Dps proteins (NpDps1-5). Our previous studies have indicated physiological differences, as well as cell-specific expression, among these five proteins. Three of the five NpDps proteins, NpDps1, -2, and -3, were classified as canonical Dps proteins. To further investigate their properties and possible importance for physiological function, here we characterized and compared them in vitro Nondenaturing PAGE, gel filtration, and dynamic light-scattering experiments disclosed that the three NpDps proteins exist as multimeric protein species in the bacterial cell. We also demonstrate Dps-mediated iron oxidation catalysis in the presence of H2O2 However, no iron oxidation with O2 as the electron acceptor was detected under our experimental conditions. In modeled structures of NpDps1, -2, and -3, protein channels were identified that could serve as the entrance for ferrous iron into the dodecameric structures. Furthermore, we could demonstrate pH-dependent DNA-binding properties for NpDps2 and -3. This study adds critical insights into the functions and stabilities of the three canonical Dps proteins from N. punctiforme and suggests that each of the Dps proteins within this bacterium has a specific biochemical property and function.
Keywords: DNA binding protein; DNA-binding proteins from starved cells; bacterial metabolism; cyanobacteria; ferritin; iron; metal homeostasis; multifunctional protein; oxidative stress; reactive oxygen species (ROS).
© 2018 Howe et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
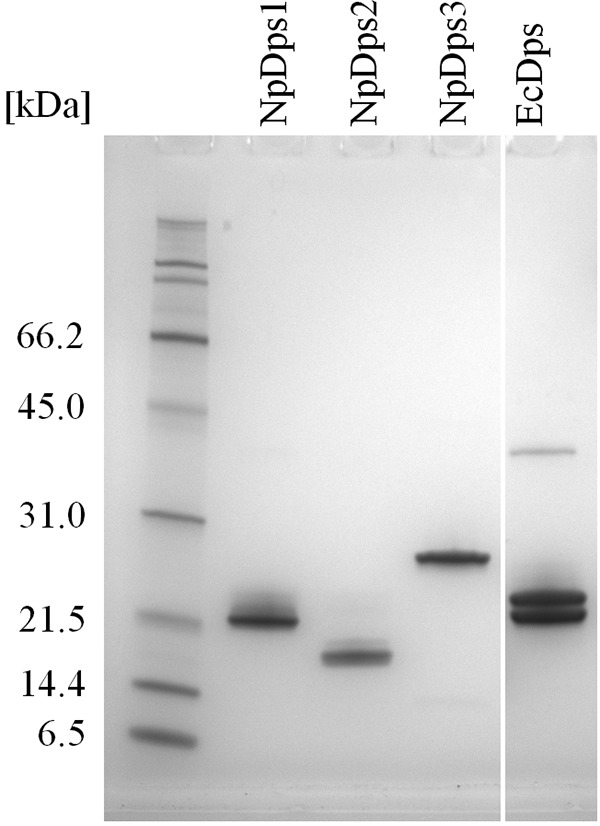
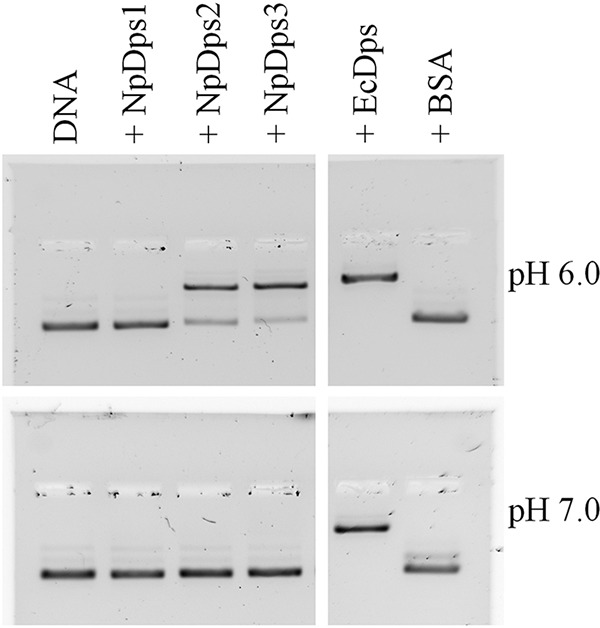
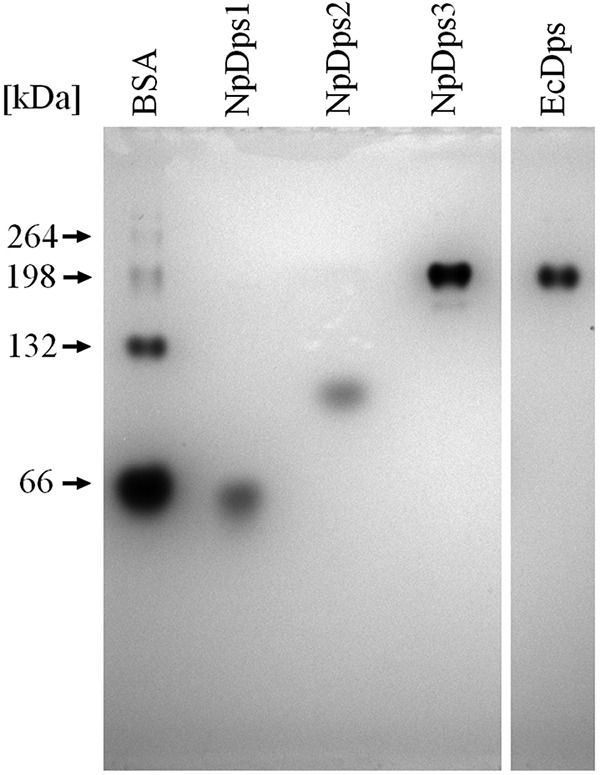
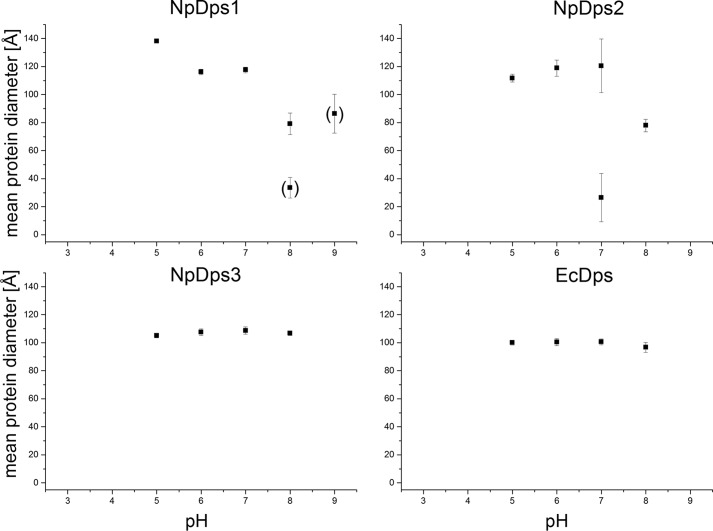
References
-
- Zhao G., Ceci P., Ilari A., Giangiacomo L., Laue T. M., Chiancone E., and Chasteen N. D. (2002) Iron and hydrogen peroxide detoxification properties of DNA-binding protein from starved cells. A “ferritin-like” DNA-binding protein of Escherichia coli. J. Biol. Chem. 277, 27689–27696 10.1074/.M202094200 - DOI - PubMed
-
- Calhoun L. N., and Kwon Y. M. (2011) Structure, function and regulation of the DNA-binding protein Dps and its role in acid and oxidative stress resistance in Escherichia coli: a review. J. Appl. Microbiol. 110, 375–386 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
