

Strong succession in arbuscular mycorrhizal fungal communities
- PMID: 30171254
- PMCID: PMC6298956
- DOI: 10.1038/s41396-018-0264-0
Strong succession in arbuscular mycorrhizal fungal communities
Abstract
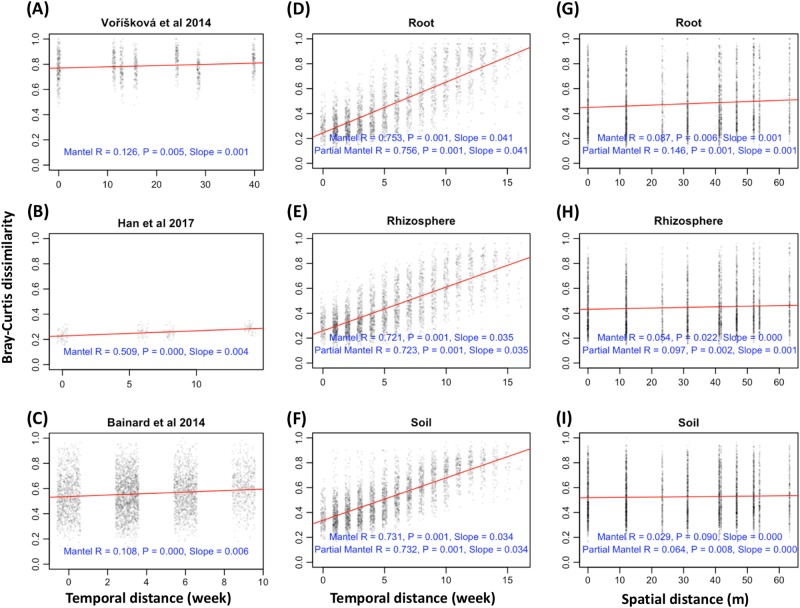
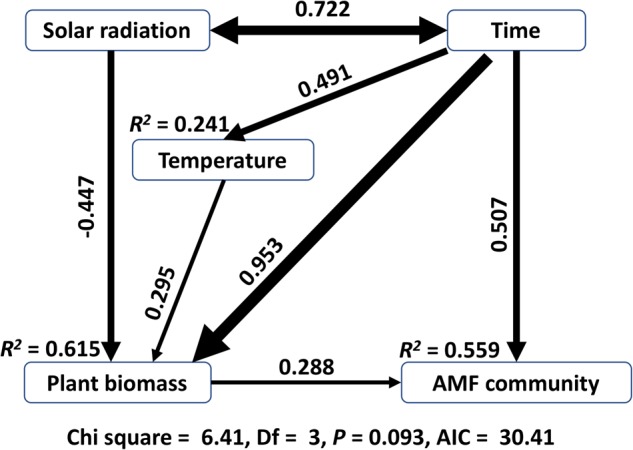
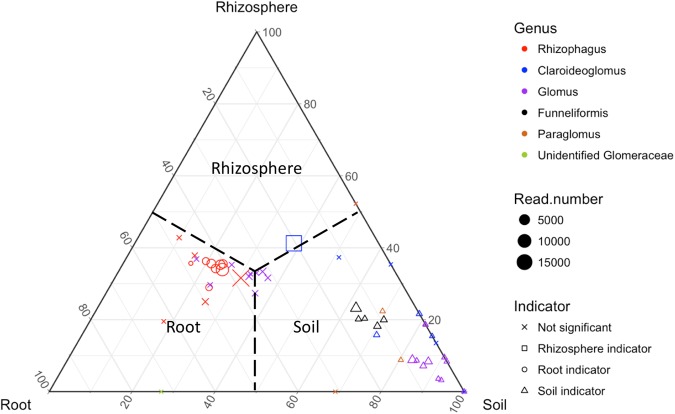
The ecology of fungi lags behind that of plants and animals because most fungi are microscopic and hidden in their substrates. Here, we address the basic ecological process of fungal succession in nature using the microscopic, arbuscular mycorrhizal fungi (AMF) that form essential mutualisms with 70-90% of plants. We find a signal for temporal change in AMF community similarity that is 40-fold stronger than seen in the most recent studies, likely due to weekly samplings of roots, rhizosphere and soil throughout the 17 weeks from seedling to fruit maturity and the use of the fungal DNA barcode to recognize species in a simple, agricultural environment. We demonstrate the patterns of nestedness and turnover and the microbial equivalents of the processes of immigration and extinction, that is, appearance and disappearance. We also provide the first evidence that AMF species co-exist rather than simply co-occur by demonstrating negative, density-dependent population growth for multiple species. Our study shows the advantages of using fungi to test basic ecological hypotheses (e.g., nestedness v. turnover, immigration v. extinction, and coexistence theory) over periods as short as one season.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




References
-
- Smith SE, Read DJ (2010). Mycorrhizal symbiosis, 3rd edn. Academic Press: London.
-
- Taylor JD, Helgason T, Öpik M (2017). Molecular Community Ecology of Arbuscular Mycorrhizal Fungi. Taylor JD, Helgason T, Öpik M (2017) Molecular community Ecology of Arbuscular Mycorrhizal Fungi. In: Dighton J, White JF, eds. The Fungal Community: its Organization and Role in the Ecosystem, 4th edn. CRC Press, pp 1–26.
-
- Rillig MC. Arbuscular mycorrhizae and terrestrial ecosystem processes. Ecol Lett. 2004;7:740–54. doi: 10.1111/j.1461-0248.2004.00620.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
