

Mechanism of loading and translocation of type VI secretion system effector Tse6
- PMID: 30177742
- PMCID: PMC6488228
- DOI: 10.1038/s41564-018-0238-z
Mechanism of loading and translocation of type VI secretion system effector Tse6
Abstract
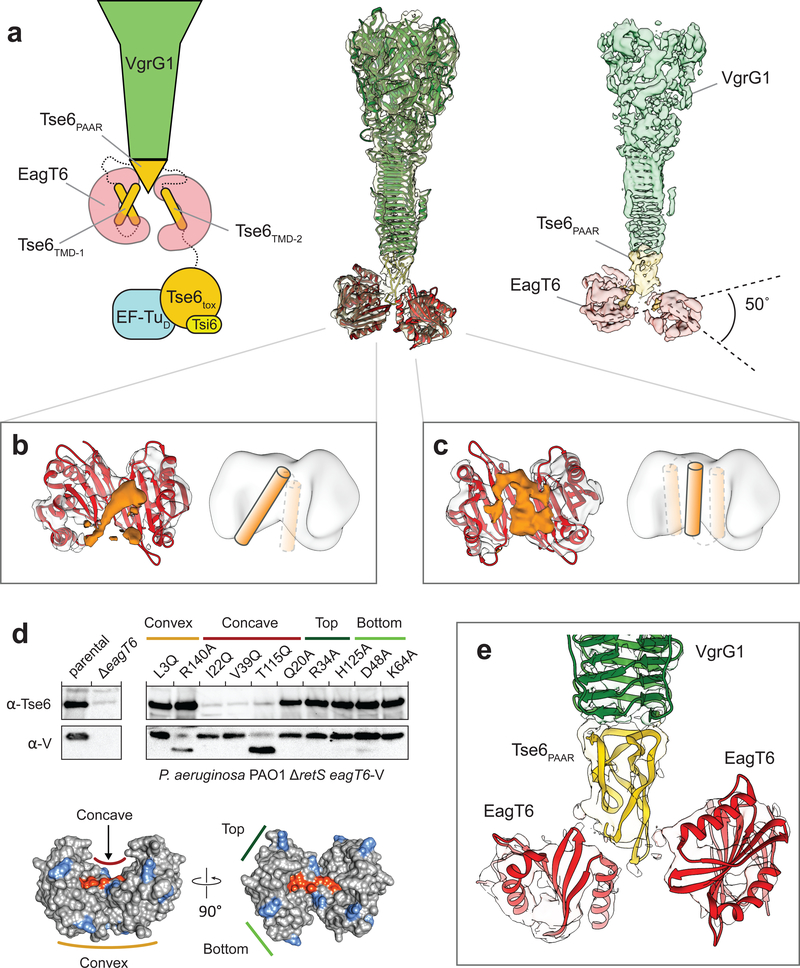
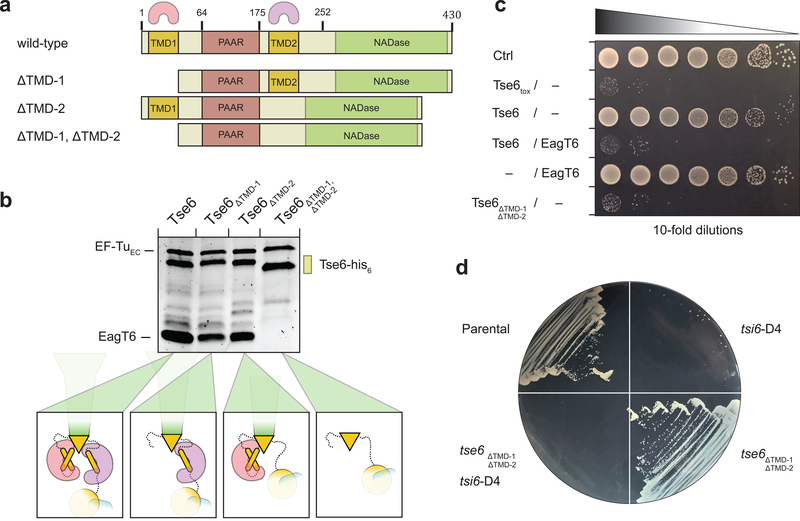
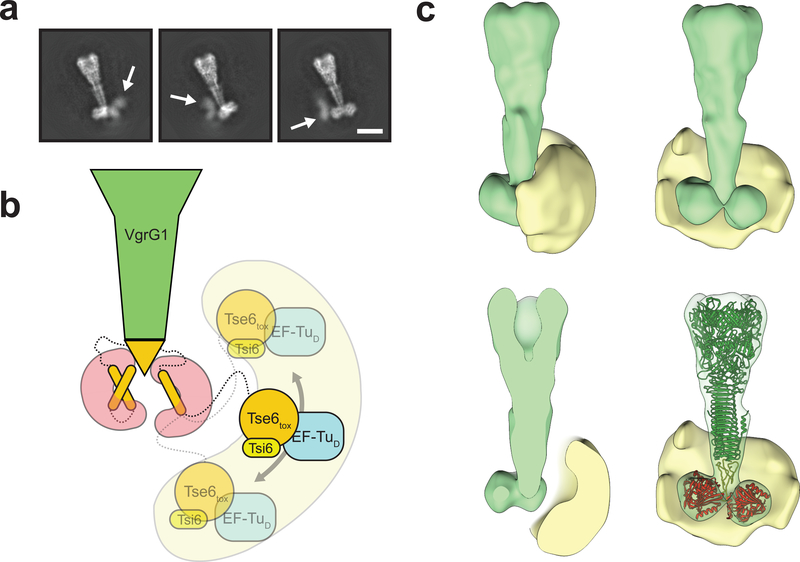
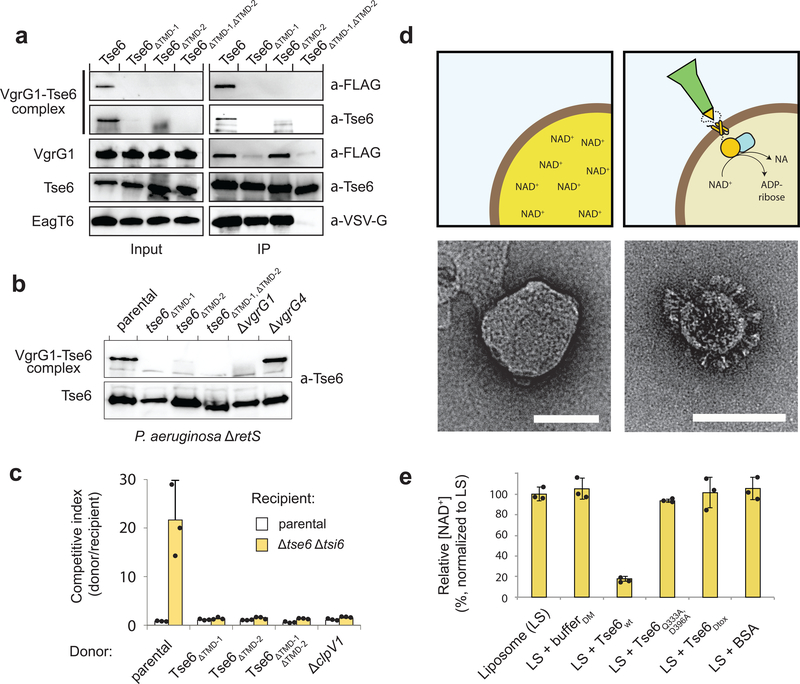
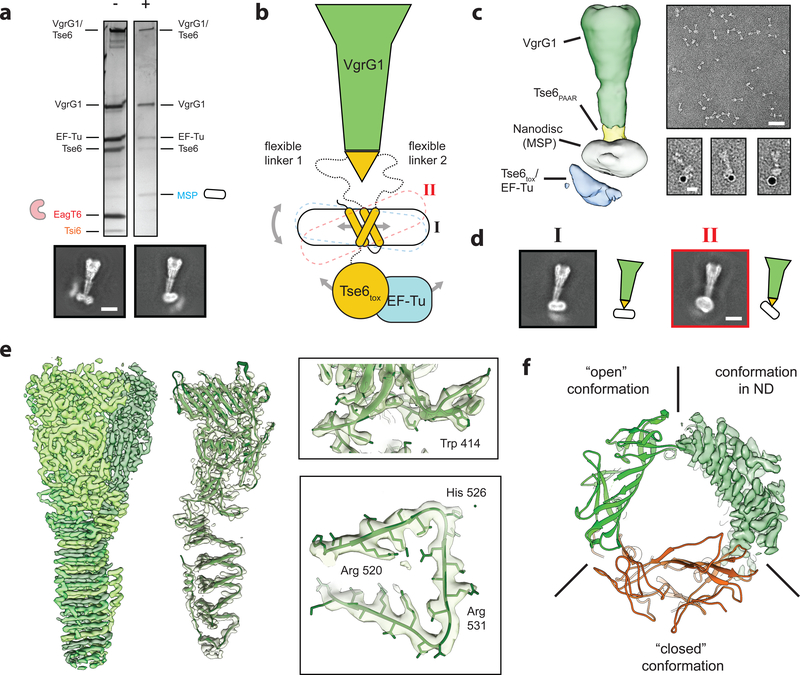
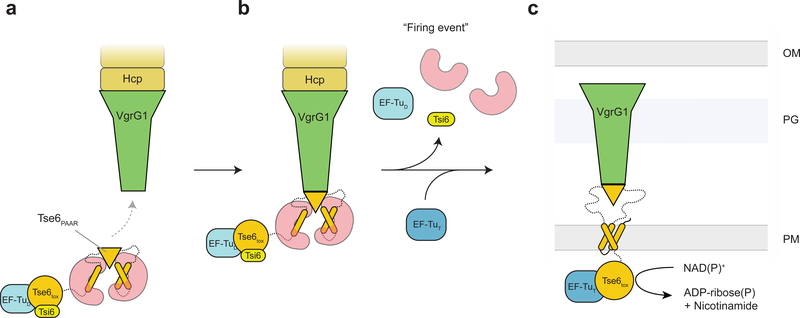
The type VI secretion system (T6SS) primarily functions to mediate antagonistic interactions between contacting bacterial cells, but also mediates interactions with eukaryotic hosts. This molecular machine secretes antibacterial effector proteins by undergoing cycles of extension and contraction; however, how effectors are loaded into the T6SS and subsequently delivered to target bacteria remains poorly understood. Here, using electron cryomicroscopy, we analysed the structures of the Pseudomonas aeruginosa effector Tse6 loaded onto the T6SS spike protein VgrG1 in solution and embedded in lipid nanodiscs. In the absence of membranes, Tse6 stability requires the chaperone EagT6, two dimers of which interact with the hydrophobic transmembrane domains of Tse6. EagT6 is not directly involved in Tse6 delivery but is crucial for its loading onto VgrG1. VgrG1-loaded Tse6 spontaneously enters membranes and its toxin domain translocates across a lipid bilayer, indicating that effector delivery by the T6SS does not require puncturing of the target cell inner membrane by VgrG1. Eag chaperone family members from diverse Proteobacteria are often encoded adjacent to putative toxins with predicted transmembrane domains and we therefore anticipate that our findings will be generalizable to numerous T6SS-exported membrane-associated effectors.
Conflict of interest statement
Competing interests
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
