

Interchromosomal interactions: A genomic love story of kissing chromosomes
- PMID: 30181316
- PMCID: PMC6314556
- DOI: 10.1083/jcb.201806052
Interchromosomal interactions: A genomic love story of kissing chromosomes
Abstract
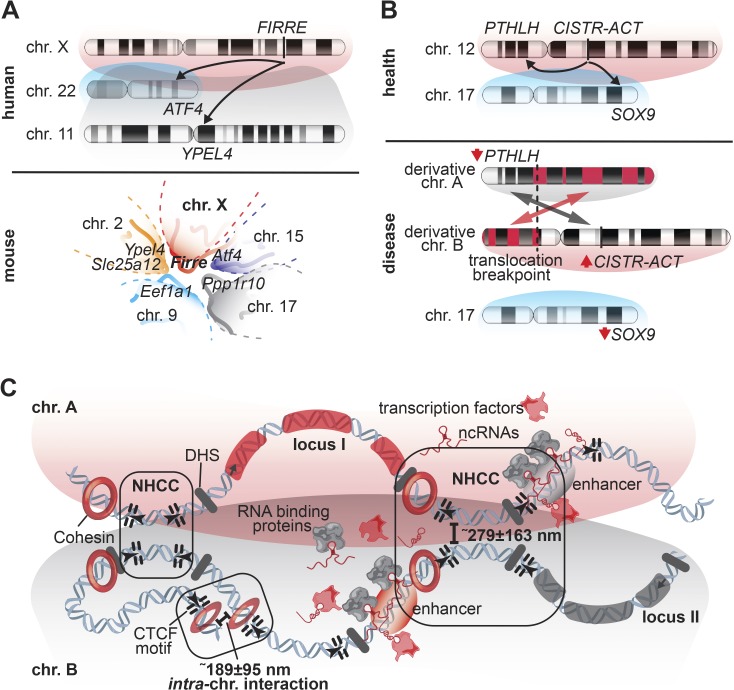
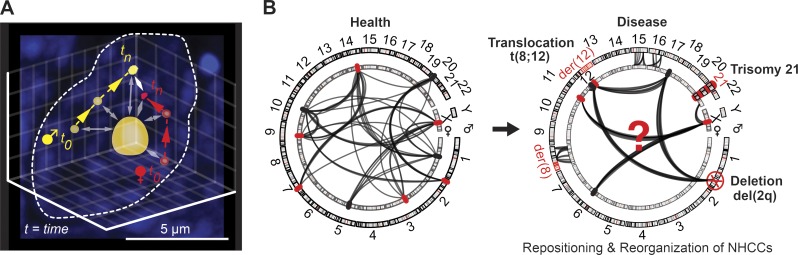
Nuclei require a precise three- and four-dimensional organization of DNA to establish cell-specific gene-expression programs. Underscoring the importance of DNA topology, alterations to the nuclear architecture can perturb gene expression and result in disease states. More recently, it has become clear that not only intrachromosomal interactions, but also interchromosomal interactions, a less studied feature of chromosomes, are required for proper physiological gene-expression programs. Here, we review recent studies with emerging insights into where and why cross-chromosomal communication is relevant. Specifically, we discuss how long noncoding RNAs (lncRNAs) and three-dimensional gene positioning are involved in genome organization and how low-throughput (live-cell imaging) and high-throughput (Hi-C and SPRITE) techniques contribute to understand the fundamental properties of interchromosomal interactions.
© 2018 Maass et al.
Figures

References
-
- Allahyar A., Vermeulen C., Bouwman B.A.M., Krijger P.H.L., Verstegen M.J.A.M., Geeven G., van Kranenburg M., Pieterse M., Straver R., Haarhuis J.H.I., et al. . 2018. Enhancer hubs and loop collisions identified from single-allele topologies. Nat. Genet. 50:1151–1160. 10.1038/s41588-018-0161-5 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
