

Effects of PERK eIF2α Kinase Inhibitor against Toxoplasma gondii
- PMID: 30181373
- PMCID: PMC6201070
- DOI: 10.1128/AAC.01442-18
Effects of PERK eIF2α Kinase Inhibitor against Toxoplasma gondii
Abstract
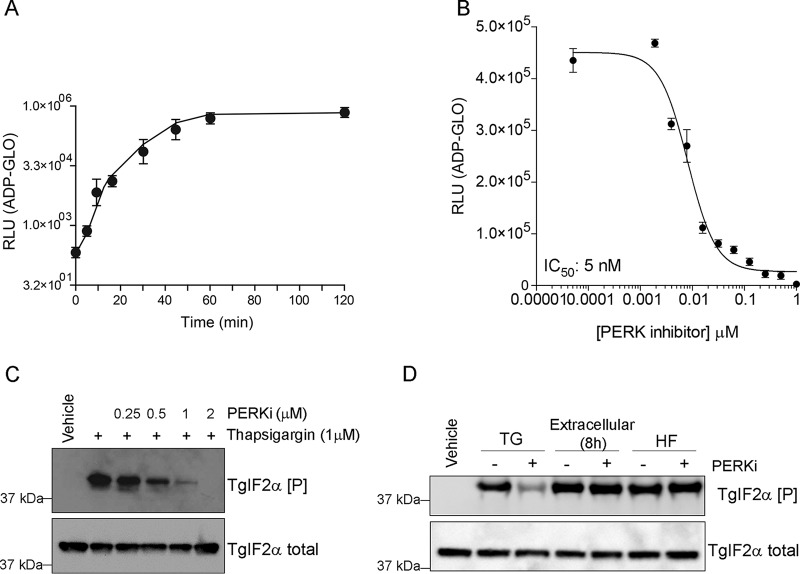
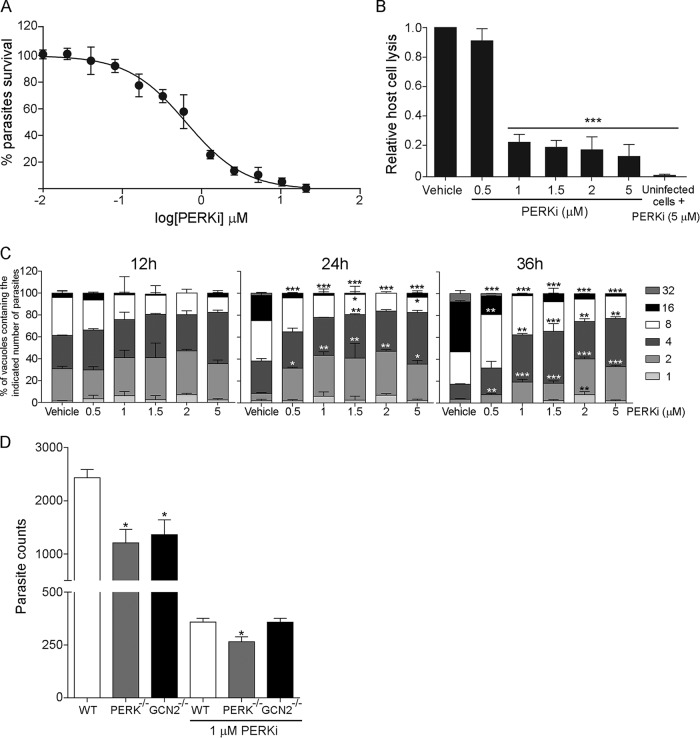
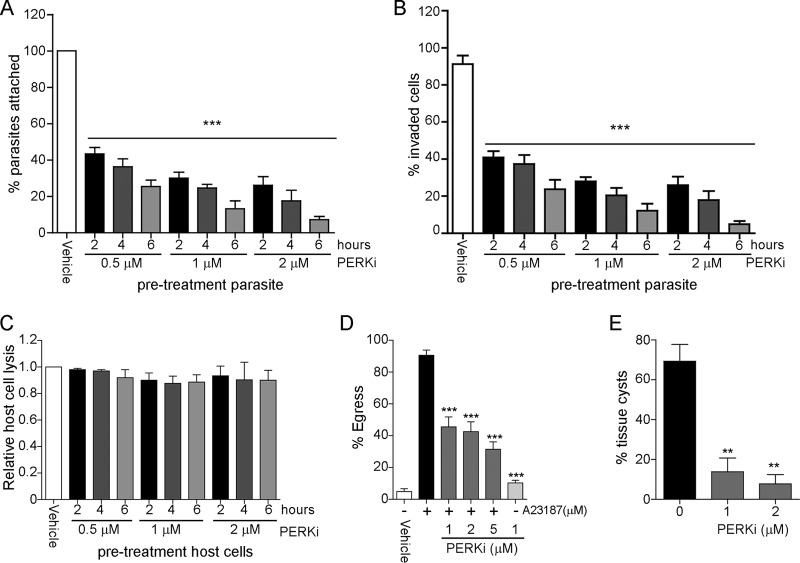
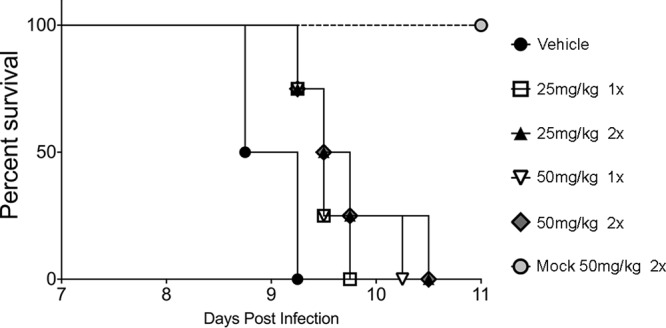
Toxoplasma gondii is an obligate intracellular parasite that has infected one-third of the population. Upon infection of warm-blooded vertebrates, the replicating form of the parasite (tachyzoite) converts into a latent form (bradyzoite) present in tissue cysts. During immune deficiency, bradyzoites can reconvert into tachyzoites and cause life-threatening toxoplasmosis. We previously reported that translational control through phosphorylation of the α subunit of T. gondii eukaryotic initiation factor 2 (eIF2α) (TgIF2α) is a critical component of the parasite stress response. Diverse stresses can induce the conversion of tachyzoites to bradyzoites, including those disrupting the parasite's endoplasmic reticulum (ER) (ER stress). Toxoplasma possesses four eIF2α kinases, one of which (TgIF2K-A) localizes to the parasite ER analogously to protein kinase R-like endoplasmic reticulum kinase (PERK), the eIF2α kinase that responds to ER stress in mammalian cells. Here, we investigated the effects of a PERK inhibitor (PERKi) on Toxoplasma Our results show that the PERKi GSK2606414 blocks the enzymatic activity of TgIF2K-A and reduces TgIF2α phosphorylation specifically in response to ER stress. PERKi also significantly impeded multiple steps of the tachyzoite lytic cycle and sharply lowered the frequency of bradyzoite differentiation in vitro Pretreatment of host cells with PERKi prior to infection did not affect parasite infectivity, and PERKi still impaired parasite replication in host cells lacking PERK. In mice, PERKi conferred modest protection from a lethal dose of Toxoplasma Our findings represent the first pharmacological evidence supporting TgIF2K-A as an attractive new target for the treatment of toxoplasmosis.
Keywords: Apicomplexa; antiparasitics; drugs; parasite; protozoa; stress response; translation.
Copyright © 2018 American Society for Microbiology.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
