

Cochaperones enable Hsp70 to use ATP energy to stabilize native proteins out of the folding equilibrium
- PMID: 30181618
- PMCID: PMC6123477
- DOI: 10.1038/s41598-018-31641-w
Cochaperones enable Hsp70 to use ATP energy to stabilize native proteins out of the folding equilibrium
Abstract
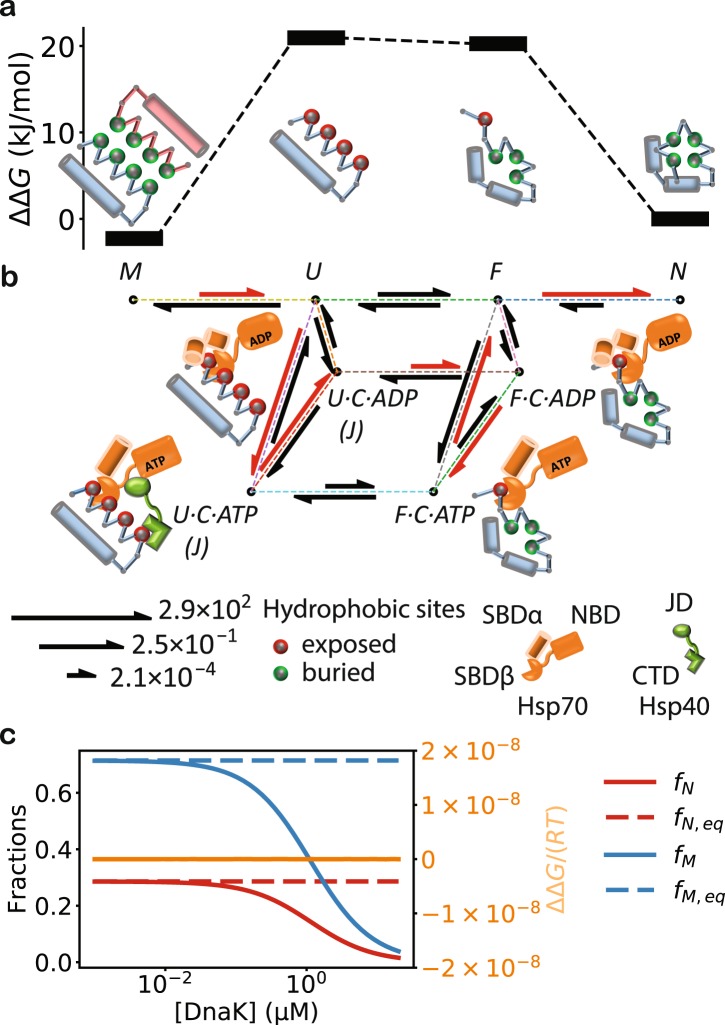
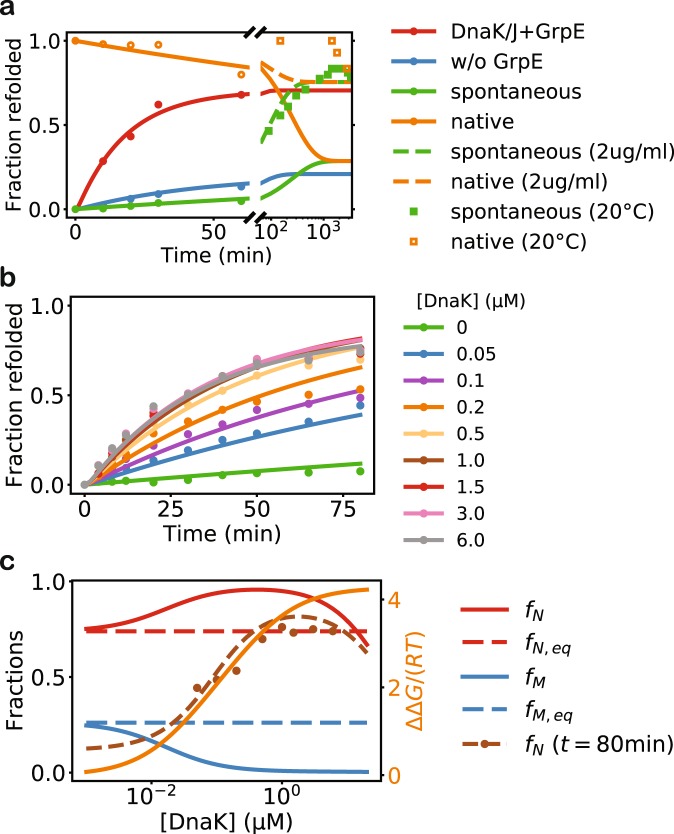
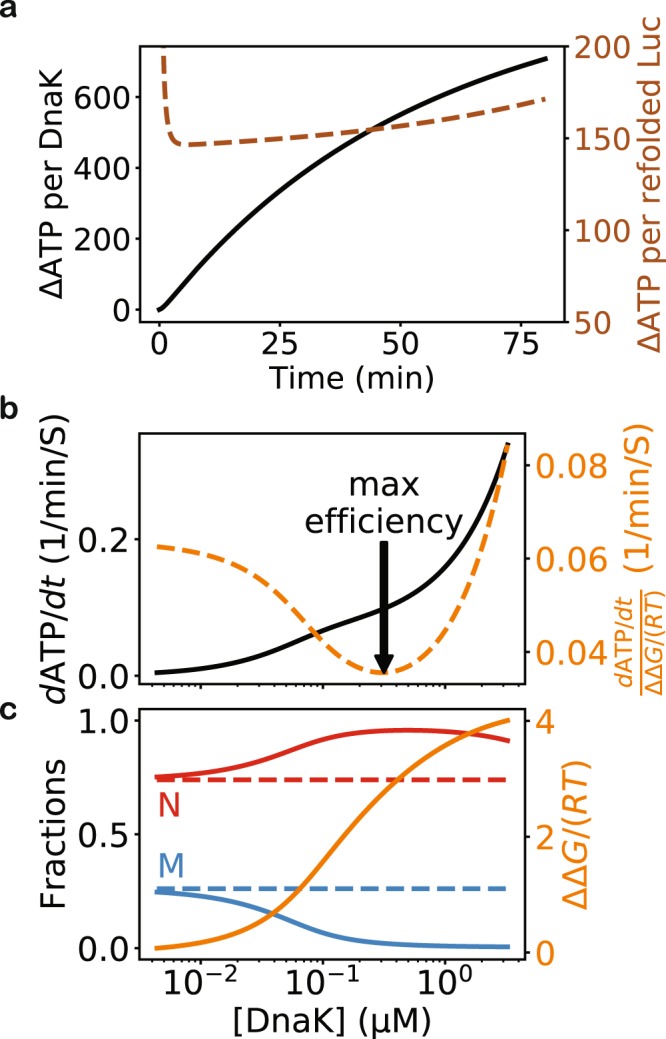
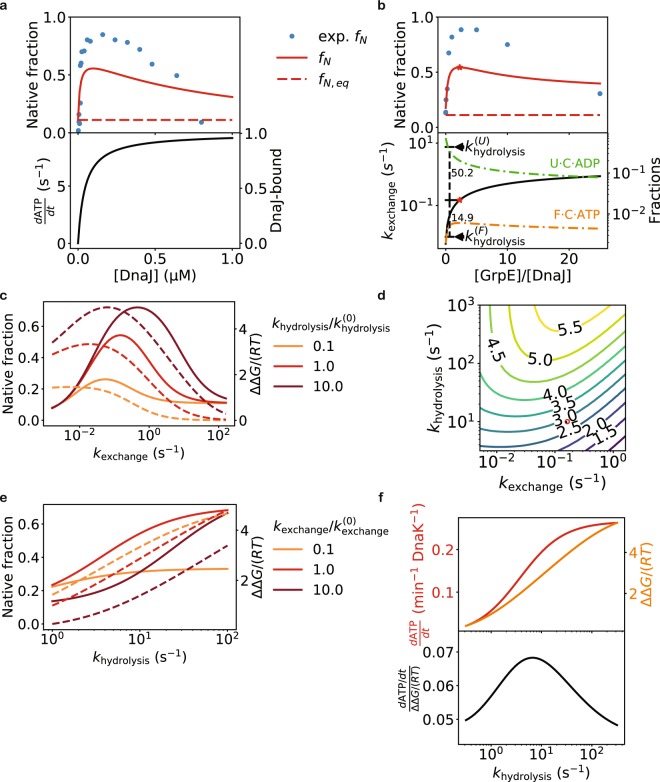
The heat shock protein 70 (Hsp70) chaperones, vital to the proper folding of proteins inside cells, consume ATP and require cochaperones in assisting protein folding. It is unclear whether Hsp70 can utilize the free energy from ATP hydrolysis to fold a protein into a native state that is thermodynamically unstable in the chaperone-free equilibrium. Here I present a model of Hsp70-mediated protein folding, which predicts that Hsp70, as a result of differential stimulation of ATP hydrolysis by its Hsp40 cochaperone, dissociates faster from a substrate in fold-competent conformations than from one in misfolding-prone conformations, thus elevating the native concentration above and suppressing the misfolded concentration below their respective equilibrium values. Previous models would not make or imply these predictions, which are experimentally testable. My model quantitatively reproduces experimental refolding kinetics, predicts how modulations of the Hsp70/Hsp40 chaperone system affect protein folding, and suggests new approaches to regulating cellular protein quality.
Conflict of interest statement
The author declares no competing interests.
Figures


Similar articles
-
Non-Equilibrium Protein Folding and Activation by ATP-Driven Chaperones.Biomolecules. 2022 Jun 15;12(6):832. doi: 10.3390/biom12060832. Biomolecules. 2022. PMID: 35740957 Free PMC article.
-
Protein folding rates and thermodynamic stability are key determinants for interaction with the Hsp70 chaperone system.Protein Sci. 2012 Oct;21(10):1489-502. doi: 10.1002/pro.2139. Protein Sci. 2012. PMID: 22886941 Free PMC article.
-
Speed-Energy-Efficiency Trade-off in Hsp70 Chaperone System.J Phys Chem B. 2024 Dec 12;128(49):12101-12113. doi: 10.1021/acs.jpcb.4c06594. Epub 2024 Dec 2. J Phys Chem B. 2024. PMID: 39622490
-
Specification of Hsp70 Function by Hsp40 Co-chaperones.Subcell Biochem. 2023;101:127-139. doi: 10.1007/978-3-031-14740-1_4. Subcell Biochem. 2023. PMID: 36520305 Review.
-
Specification of Hsp70 function by Type I and Type II Hsp40.Subcell Biochem. 2015;78:91-102. doi: 10.1007/978-3-319-11731-7_4. Subcell Biochem. 2015. PMID: 25487017 Review.
Cited by
-
Regulating glycolysis and heat shock proteins in Gannan yaks (Bos grunniens) in response to hypoxia of the Qinghai-Tibet Plateau.Arch Anim Breed. 2021 Aug 19;64(2):345-353. doi: 10.5194/aab-64-345-2021. eCollection 2021. Arch Anim Breed. 2021. PMID: 34458561 Free PMC article.
-
Proteinopathies: Deciphering Physiology and Mechanisms to Develop Effective Therapies for Neurodegenerative Diseases.Mol Neurobiol. 2022 Dec;59(12):7513-7540. doi: 10.1007/s12035-022-03042-8. Epub 2022 Oct 7. Mol Neurobiol. 2022. PMID: 36205914 Review.
-
Misfolding and aggregation in neurodegenerative diseases: protein quality control machinery as potential therapeutic clearance pathways.Cell Commun Signal. 2024 Aug 30;22(1):421. doi: 10.1186/s12964-024-01791-8. Cell Commun Signal. 2024. PMID: 39215343 Free PMC article. Review.
-
Non-Equilibrium Protein Folding and Activation by ATP-Driven Chaperones.Biomolecules. 2022 Jun 15;12(6):832. doi: 10.3390/biom12060832. Biomolecules. 2022. PMID: 35740957 Free PMC article.
-
ATP-Driven Nonequilibrium Activation of Kinase Clients by the Molecular Chaperone Hsp90.Biophys J. 2020 Oct 20;119(8):1538-1549. doi: 10.1016/j.bpj.2020.08.038. Epub 2020 Sep 11. Biophys J. 2020. PMID: 33038305 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
