

Glycolytic Stimulation Is Not a Requirement for M2 Macrophage Differentiation
- PMID: 30184486
- PMCID: PMC6449248
- DOI: 10.1016/j.cmet.2018.08.012
Glycolytic Stimulation Is Not a Requirement for M2 Macrophage Differentiation
Abstract
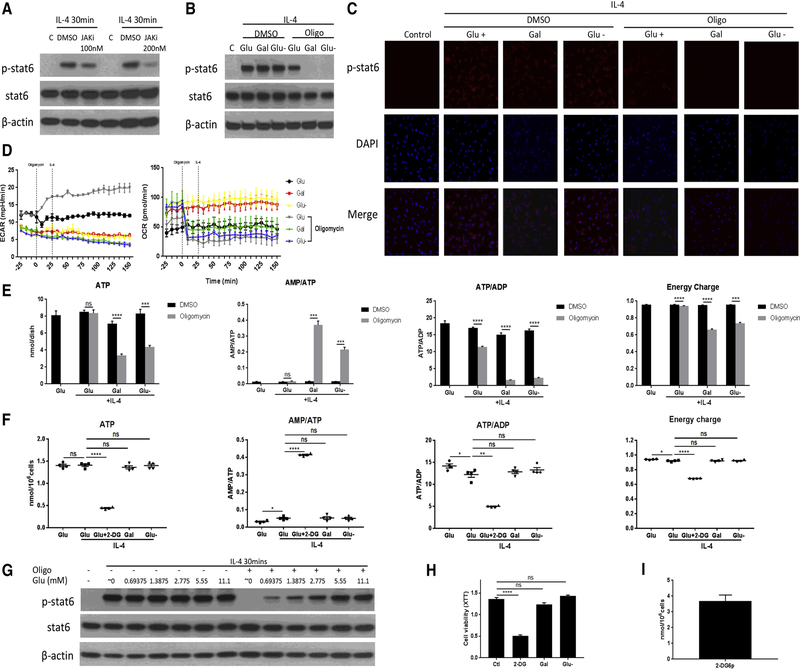
Enhanced glucose uptake and a switch to glycolysis are key traits of M1 macrophages, whereas enhanced fatty acid oxidation and oxidative phosphorylation are the main metabolic characteristics of M2 macrophages. Recent studies challenge this traditional view, indicating that glycolysis may also be critically important for M2 macrophage differentiation, based on experiments with 2-DG. Here we confirm the inhibitory effect of 2-DG on glycolysis, but also demonstrate that 2-DG impairs oxidative phosphorylation and significantly reduces 13C-labeled Krebs cycle metabolites and intracellular ATP levels. These metabolic derangements were associated with reduced JAK-STAT6 pathway activity and M2 differentiation marker expression. While glucose deprivation and glucose substitution with galactose effectively suppressed glycolytic activity, there was no effective suppression of oxidative phosphorylation, intracellular ATP levels, STAT6 phosphorylation, and M2 differentiation marker expression. These data indicate that glycolytic stimulation is not required for M2 macrophage differentiation as long as oxidative phosphorylation remains active.
Keywords: 2-DG; M2 macrophage; alternative stimulation; glucose; glycolysis; interleukin-4; metabolism.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
None for any author.
Figures






References
-
- Akira S, Uematsu S, and Takeuchi O (2006). Pathogen recognition and innate immunity. Cell 124, 783–801. - PubMed
-
- Biswas SK and Mantovani A (2010). Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nature Immunology 11, 889–896. - PubMed
-
- Burelle Y, Lamoureux MC, Péronnet F, Massicotte D, Lavoie C. (2006) Comparison of exogenous glucose, fructose and galactose oxidation during exercise using 13C-labelling. Br J Nutr 96:56–61. - PubMed
-
- Chen W, Hoerter J, Guéron M (1996) A comparison of AMP degradation in the perfused rat heart during 2-deoxy-D-glucose perfusion and anoxia. Part I: The release of adenosine and inosine. J Mol Cell Cardiol 28:2163–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
