

A fluid-to-solid jamming transition underlies vertebrate body axis elongation
- PMID: 30185907
- PMCID: PMC6148385
- DOI: 10.1038/s41586-018-0479-2
A fluid-to-solid jamming transition underlies vertebrate body axis elongation
Abstract
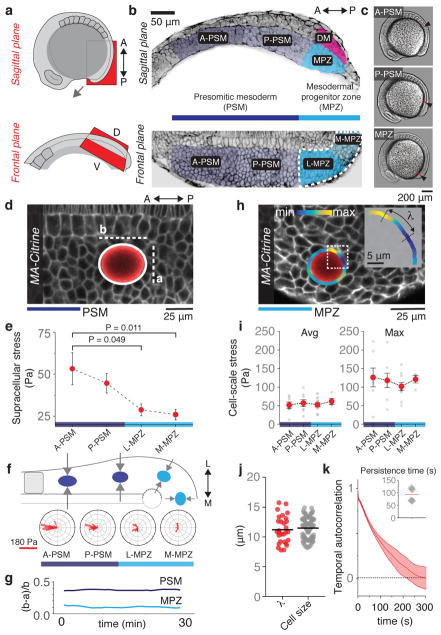
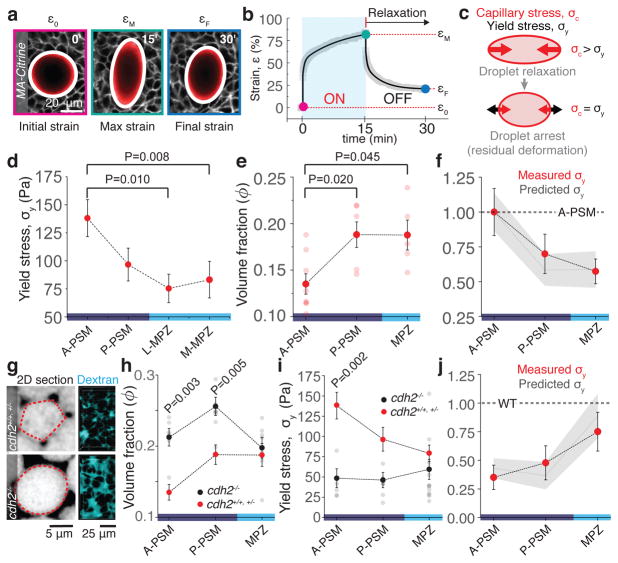
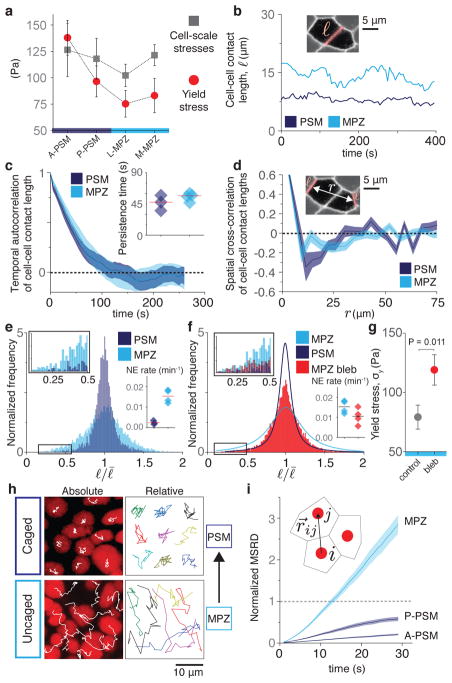
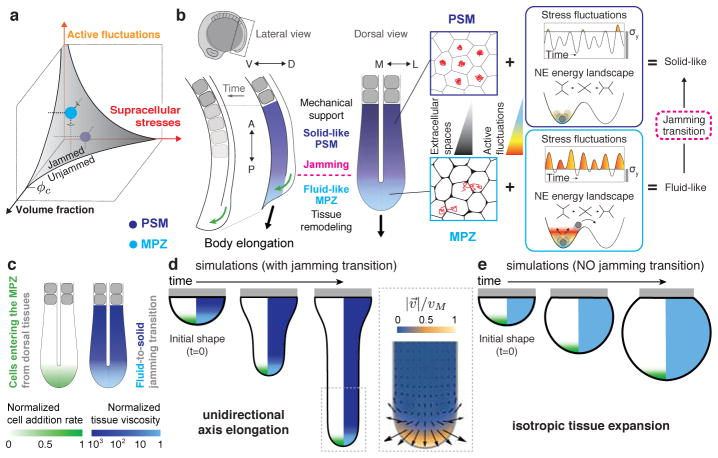
Just as in clay moulding or glass blowing, physically sculpting biological structures requires the constituent material to locally flow like a fluid while maintaining overall mechanical integrity like a solid. Disordered soft materials, such as foams, emulsions and colloidal suspensions, switch from fluid-like to solid-like behaviours at a jamming transition1-4. Similarly, cell collectives have been shown to display glassy dynamics in 2D and 3D5,6 and jamming in cultured epithelial monolayers7,8, behaviours recently predicted theoretically9-11 and proposed to influence asthma pathobiology8 and tumour progression12. However, little is known about whether these seemingly universal behaviours occur in vivo13 and, specifically, whether they play any functional part during embryonic morphogenesis. Here, by combining direct in vivo measurements of tissue mechanics with analysis of cellular dynamics, we show that during vertebrate body axis elongation, posterior tissues undergo a jamming transition from a fluid-like behaviour at the extending end, the mesodermal progenitor zone, to a solid-like behaviour in the presomitic mesoderm. We uncover an anteroposterior, N-cadherin-dependent gradient in yield stress that provides increasing mechanical integrity to the presomitic mesoderm, consistent with the tissue transiting from a wetter to a dryer foam-like architecture. Our results show that cell-scale stresses fluctuate rapidly (within about 1 min), enabling cell rearrangements and effectively 'melting' the tissue at the growing end. Persistent (more than 0.5 h) stresses at supracellular scales, rather than cell-scale stresses, guide morphogenetic flows in fluid-like tissue regions. Unidirectional axis extension is sustained by the reported rigidification of the presomitic mesoderm, which mechanically supports posterior, fluid-like tissues during remodelling before their maturation. The spatiotemporal control of fluid-like and solid-like tissue states may represent a generic physical mechanism of embryonic morphogenesis.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
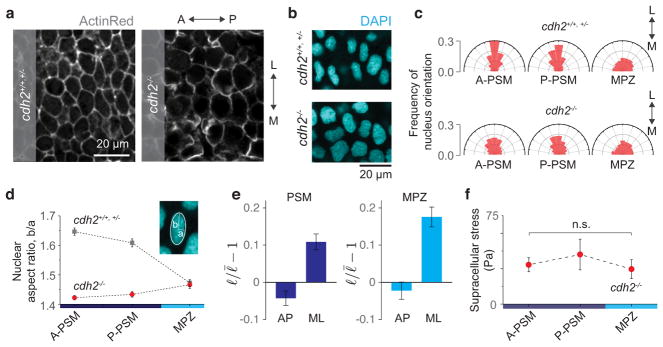
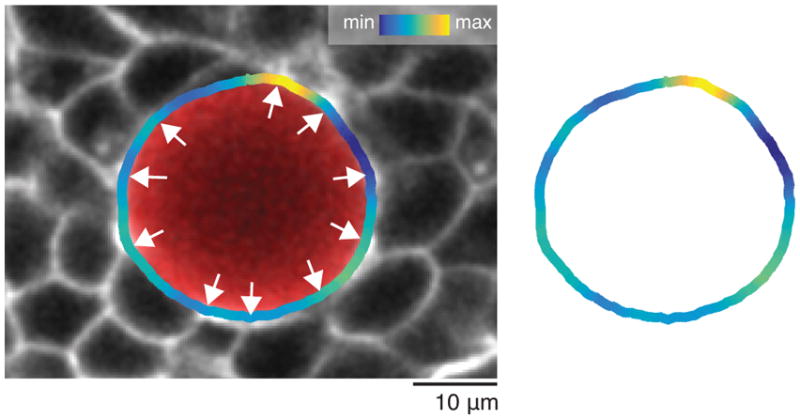

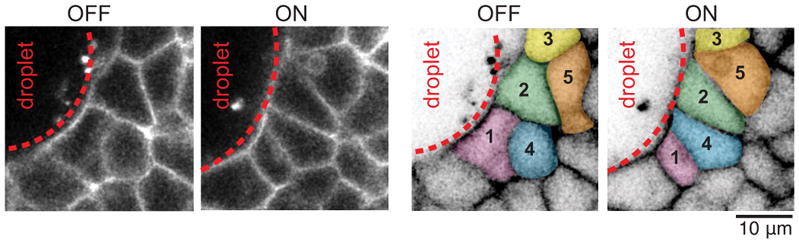
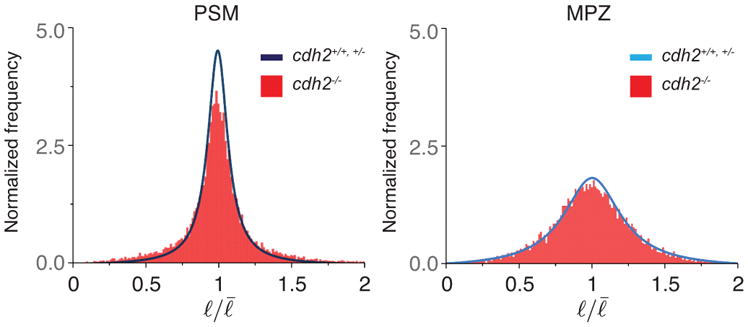
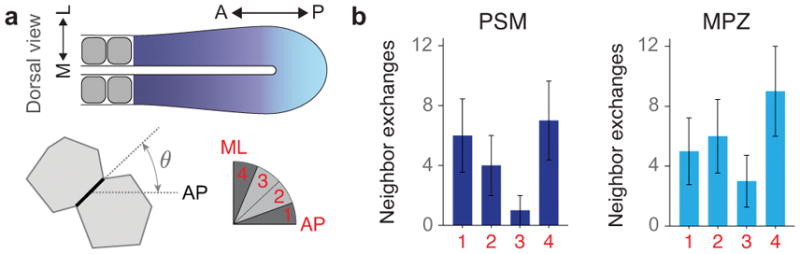

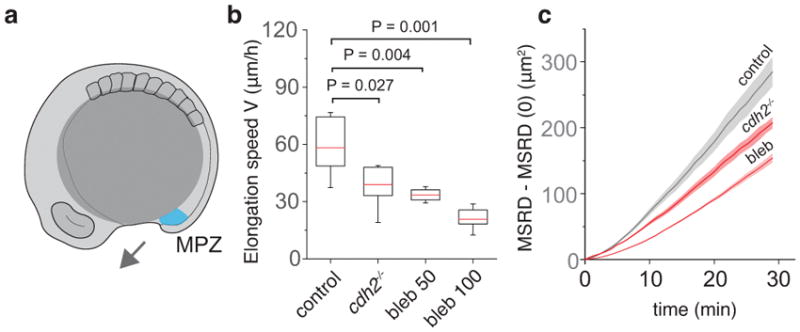
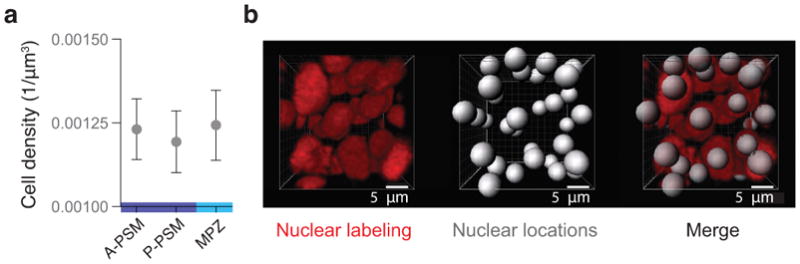
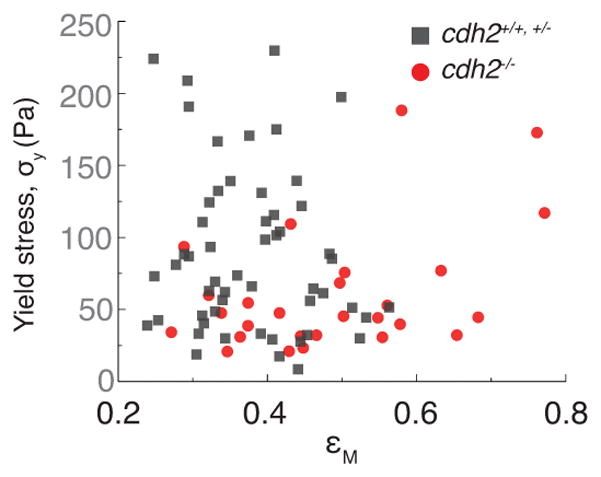
Comment in
-
Tissue 'melting' sculpts embryo.Nature. 2018 Sep;561(7723):315-316. doi: 10.1038/d41586-018-06108-7. Nature. 2018. PMID: 30224727 No abstract available.
References
-
- Cohen-Addad S, Höhler R, Pitois O. Flow in Foams and Flowing Foams. Annu Rev Fluid Mech. 2013;45:241–267.
-
- Liu AJ, Nagel SR. Jamming is just not cool any more. Nature. 1998;396:21–22.
-
- Bonn D, Denn MM, Berthier L, Divoux T, Manneville S. Yield stress materials in soft condensed matter. Rev Mod Phys. 2017;89:15.
-
- Trappe V, Prasad V, Cipelletti L, Segre PN, Weitz DA. Jamming phase diagram for attractive particles. Nature. 2001;411:772–775. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
