

Citalopram Reduces Aggregation of ATXN3 in a YAC Transgenic Mouse Model of Machado-Joseph Disease
- PMID: 30187384
- PMCID: PMC6417975
- DOI: 10.1007/s12035-018-1331-2
Citalopram Reduces Aggregation of ATXN3 in a YAC Transgenic Mouse Model of Machado-Joseph Disease
Abstract
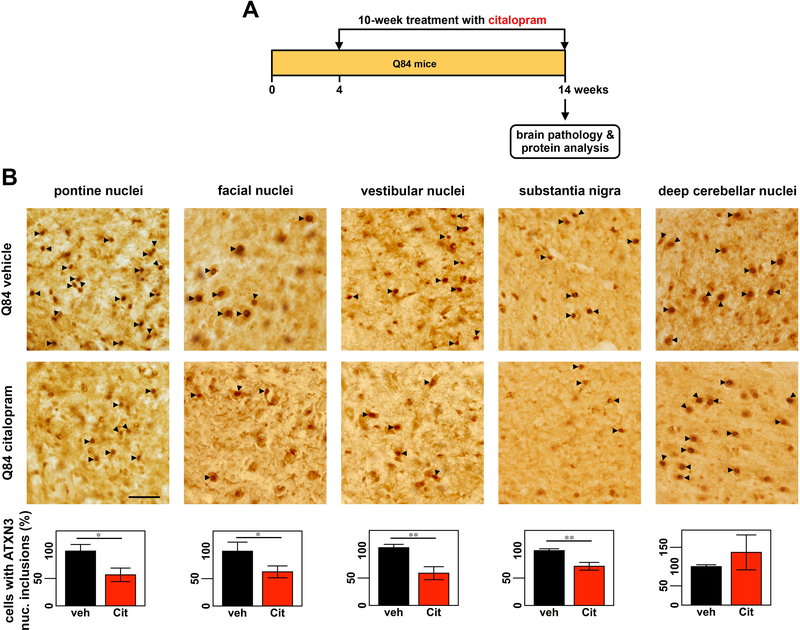
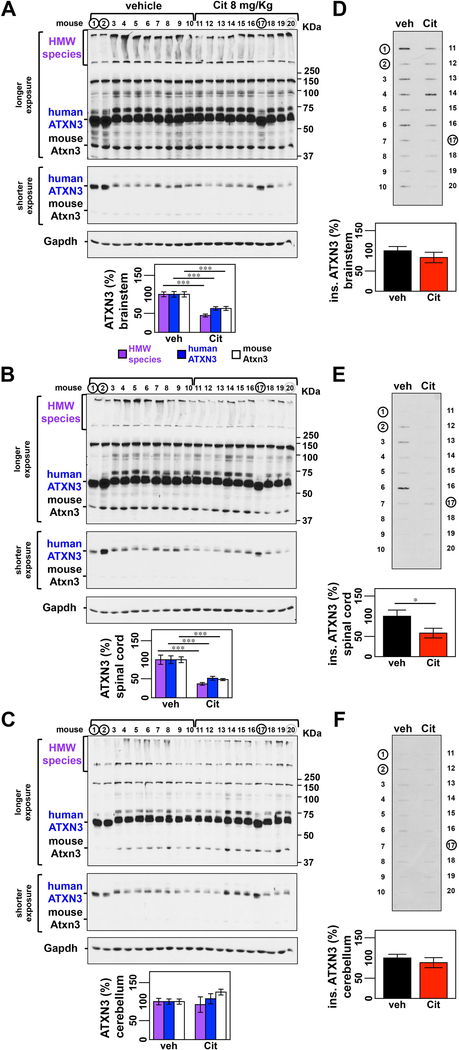
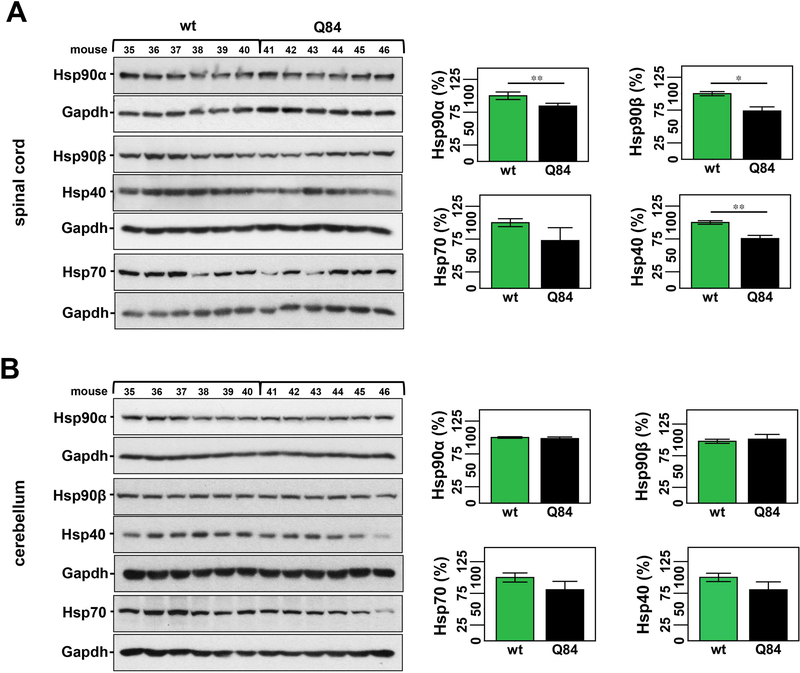
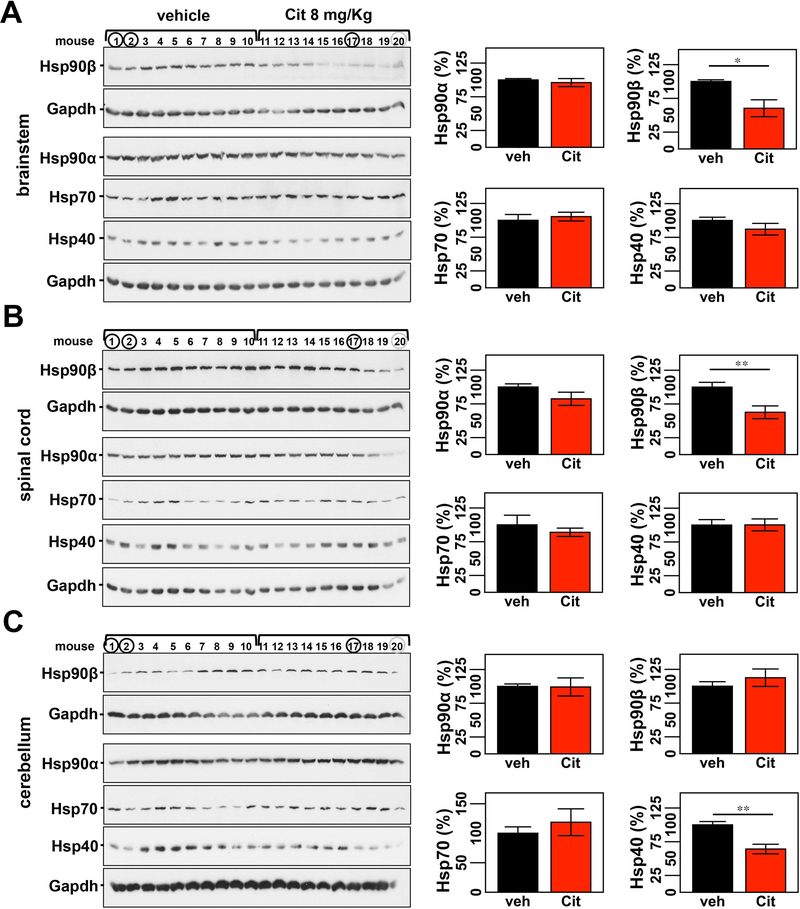
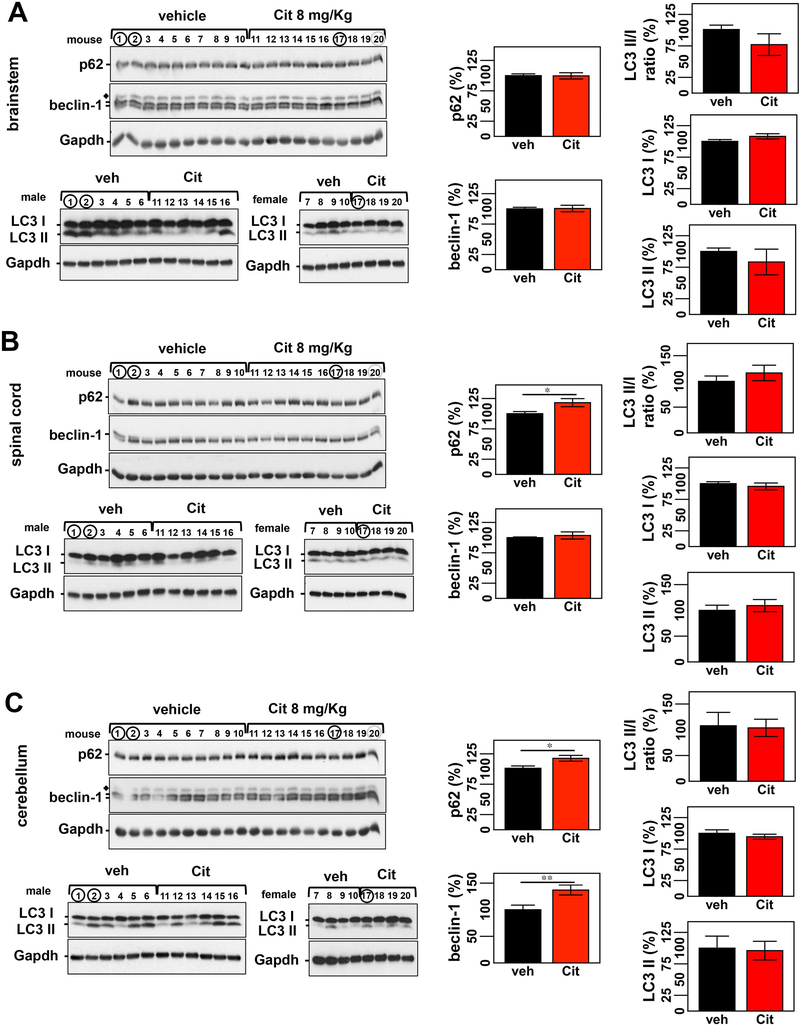
Machado-Joseph disease, also known as spinocerebellar ataxia type 3, is a fatal polyglutamine disease with no disease-modifying treatment. The selective serotonin reuptake inhibitor citalopram was shown in nematode and mouse models to be a compelling repurposing candidate for Machado-Joseph disease therapeutics. We sought to confirm the efficacy of citalopram to decrease ATXN3 aggregation in an unrelated mouse model of Machado-Joseph disease. Four-week-old YACMJD84.2 mice and non-transgenic littermates were given citalopram 8 mg/kg in drinking water or water for 10 weeks. At the end of treatment, brains were collected for biochemical and pathological analyses. Brains of citalopram-treated YACMJD84.2 mice showed an approximate 50% decrease in the percentage of cells containing ATXN3-positive inclusions in the substantia nigra and three examined brainstem nuclei compared to controls. No differences in ATXN3 inclusion load were observed in deep cerebellar nuclei of mice. Citalopram effect on ATXN3 aggregate burden was corroborated by immunoblotting analysis. While lysates from the brainstem and cervical spinal cord of citalopram-treated mice showed a decrease in all soluble forms of ATXN3 and a trend toward reduction of insoluble ATXN3, no differences in ATXN3 levels were found between cerebella of citalopram-treated and vehicle-treated mice. Citalopram treatment altered levels of select components of the cellular protein homeostatic machinery that may be expected to enhance the capacity to refold and/or degrade mutant ATXN3. The results here obtained in a second independent mouse model of Machado-Joseph disease further support citalopram as a potential drug to be repurposed for this fatal disorder.
Keywords: Neurodegeneration; Polyglutamine; Proteinopathy; Spinocerebellar ataxia; Therapy.
Figures
Similar articles
-
Preclinical Evidence Supporting Early Initiation of Citalopram Treatment in Machado-Joseph Disease.Mol Neurobiol. 2019 May;56(5):3626-3637. doi: 10.1007/s12035-018-1332-1. Epub 2018 Sep 1. Mol Neurobiol. 2019. PMID: 30173407
-
Serotonergic signalling suppresses ataxin 3 aggregation and neurotoxicity in animal models of Machado-Joseph disease.Brain. 2015 Nov;138(Pt 11):3221-37. doi: 10.1093/brain/awv262. Epub 2015 Sep 15. Brain. 2015. PMID: 26373603 Free PMC article.
-
Unbiased screen identifies aripiprazole as a modulator of abundance of the polyglutamine disease protein, ataxin-3.Brain. 2016 Nov 1;139(11):2891-2908. doi: 10.1093/brain/aww228. Brain. 2016. PMID: 27645800 Free PMC article.
-
ATXN3: a multifunctional protein involved in the polyglutamine disease spinocerebellar ataxia type 3.Expert Rev Mol Med. 2024 Sep 25;26:e19. doi: 10.1017/erm.2024.10. Expert Rev Mol Med. 2024. PMID: 39320846 Free PMC article. Review.
-
Machado-Joseph Disease: A Stress Combating Deubiquitylating Enzyme Changing Sides.Adv Exp Med Biol. 2020;1233:237-260. doi: 10.1007/978-3-030-38266-7_10. Adv Exp Med Biol. 2020. PMID: 32274760 Review.
Cited by
-
Blood neurofilament light chain levels are associated with disease progression in a transgenic SCA3 mouse model.bioRxiv [Preprint]. 2023 Mar 1:2023.02.28.530463. doi: 10.1101/2023.02.28.530463. bioRxiv. 2023. Update in: Dis Model Mech. 2023 Sep 1;16(9):dmm050144. doi: 10.1242/dmm.050144. PMID: 36909535 Free PMC article. Updated. Preprint.
-
Treatment of neurological pathology and inflammation in Machado-Joseph disease through in vivo self-assembled siRNA.Brain. 2025 Mar 6;148(3):817-832. doi: 10.1093/brain/awae304. Brain. 2025. PMID: 39315766 Free PMC article.
-
Druggable genome screen identifies new regulators of the abundance and toxicity of ATXN3, the Spinocerebellar Ataxia type 3 disease protein.Neurobiol Dis. 2020 Apr;137:104697. doi: 10.1016/j.nbd.2019.104697. Epub 2019 Nov 26. Neurobiol Dis. 2020. PMID: 31783119 Free PMC article.
-
Preclinical Evidence Supporting Early Initiation of Citalopram Treatment in Machado-Joseph Disease.Mol Neurobiol. 2019 May;56(5):3626-3637. doi: 10.1007/s12035-018-1332-1. Epub 2018 Sep 1. Mol Neurobiol. 2019. PMID: 30173407
-
Ataxia.Continuum (Minneap Minn). 2019 Aug;25(4):1036-1054. doi: 10.1212/CON.0000000000000753. Continuum (Minneap Minn). 2019. PMID: 31356292 Free PMC article. Review.
References
-
- Schols L, Bauer P, Schmidt T, Schulte T, Riess O (2004) Autosomal dominant cerebellar ataxias: clinical features, genetics, and pathogenesis. Lancet Neurol 3 (5):291–304 - PubMed
-
- Lima M, Costa MC, Montiel R, Ferro A, Santos C, Silva C, Bettencourt C, Sousa A, Sequeiros J, Coutinho P, Maciel P (2005) Population genetics of wild-type CAG repeats in the Machado-Joseph disease gene in Portugal. Hum Hered 60 (3):156–163 - PubMed
-
- Maciel P, Costa MC, Ferro A, Rousseau M, Santos CS, Gaspar C, Barros J, Rouleau GA, Coutinho P, Sequeiros J (2001) Improvement in the molecular diagnosis of Machado-Joseph disease. Arch Neurol 58 (11):1821–1827. doi:noc10138 [pii] - PubMed
MeSH terms
Substances
Grants and funding
- SFRH/BPD/102317/2014/Foundation for Science and Technology (FCT); POPH; QREN; Governo da República Portuguesa; EU/FSE
- NORTE-01-0145-FEDER-000013/Northern Portugal Regional Operational Program (NORTE 2020)
- R01NS038712/NS/NINDS NIH HHS/United States
- SFRH/BD/78388/2011/Foundation for Science and Technology (FCT); POPH; QREN; Governo da República Portuguesa; EU/FSE
- G015617/Becky Babcox Research Fund, University of Michigan
- POCI-01-0145-FEDER-007038/European Regional Development Funds (FEDER) - Competitiveness Factors Operational Programme (COMPETE); Foundation for Science and Technology (FCT)
- U01 NS104326/NS/NINDS NIH HHS/United States
- EXPL/BIM-MEC/0239/2012/Foundation for Science and Technology (FCT); Competitiveness Factors Operational Programme (COMPETE)
- POCI-01-0145-FEDER-016818 (PTDC/NEU-NMC/3648/2014)/Foundation for Science and Technology (FCT)
- PTDC/SAU-GMG/112617/2009/Foundation for Science and Technology (FCT); Competitiveness Factors Operational Programme (COMPETE)
- R01 NS038712/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
