

NFAT5 Isoform C Controls Biomechanical Stress Responses of Vascular Smooth Muscle Cells
- PMID: 30190682
- PMCID: PMC6115610
- DOI: 10.3389/fphys.2018.01190
NFAT5 Isoform C Controls Biomechanical Stress Responses of Vascular Smooth Muscle Cells
Abstract
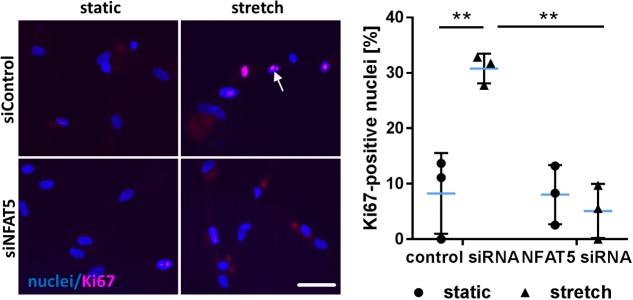
Vascular cells are continuously exposed to mechanical stress that may wreak havoc if exceeding physiological levels. Consequently, mechanisms facing such a challenge are indispensable and contribute to the adaptation of the cellular phenotype. To this end, vascular smooth muscle cells (VSMCs) activate mechanoresponsive transcription factors promoting their proliferation and migration to initiate remodeling the arterial wall. In mechanostimulated VSMCs, we identified nuclear factor of activated T-cells 5 (NFAT5) as transcriptional regulator protein and intended to unravel mechanisms controlling its expression and nuclear translocation. In cultured human VSMCs, blocking RNA synthesis diminished both baseline and stretch-induced NFAT5 mRNA expression while inhibition of the proteasome promoted accumulation of the NFAT5 protein. Detailed PCR analyses indicated a decrease in expression of NFAT5 isoform A and an increase in isoform C in mechanoactivated VSMCs. Upon overexpression, only NFAT5c was capable to enter the nucleus in control- and stretch-stimulated VSMCs. As evidenced by analyses of NFAT5c mutants, nuclear translocation required palmitoylation, phosphorylation at Y143 and was inhibited by phosphorylation at S1197. On the functional level, overexpression of NFAT5c forces its accumulation in the nucleus as well as transcriptional activity and stimulated VSMC proliferation and migration. These findings suggest that NFAT5 is continuously expressed and degraded in resting VSMCs while expression and accumulation of isoform C in the nucleus is facilitated during biomechanical stress to promote an activated VSMC phenotype.
Keywords: NFAT5; TonEBP; hypertension; transcriptional regulation; vascular smooth muscle cells; wall stress.
Figures







References
-
- Ahn J. D., Morishita R., Kaneda Y., Lee S. J., Kwon K. Y., Choi S. Y., et al. (2002). Inhibitory effects of novel AP-1 decoy oligodeoxynucleotides on vascular smooth muscle cell proliferation in vitro and neointimal formation in vivo. Circ. Res. 90 1325–1332. 10.1161/01.RES.0000023200.19316.D5 - DOI - PubMed
-
- Bergman M. R., Cheng S., Honbo N., Piacentini L., Karliner J. S., Lovett D. H. (2003). A functional activating protein 1 (AP-1) site regulates matrix metalloproteinase 2 (MMP-2) transcription by cardiac cells through interactions with JunB-Fra1 and JunB-FosB heterodimers. Biochem. J. 369 485–496. 10.1042/BJ20020707 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials