

Accelerated ex situ breeding of GBSS- and PTST1-edited cassava for modified starch
- PMID: 30191180
- PMCID: PMC6124905
- DOI: 10.1126/sciadv.aat6086
Accelerated ex situ breeding of GBSS- and PTST1-edited cassava for modified starch
Abstract
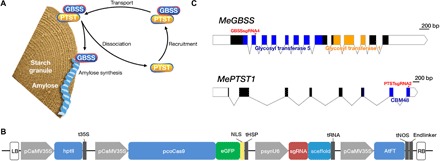
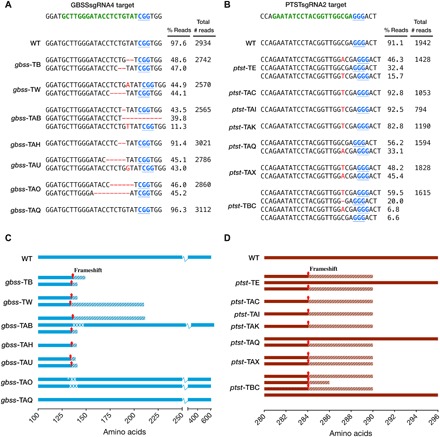
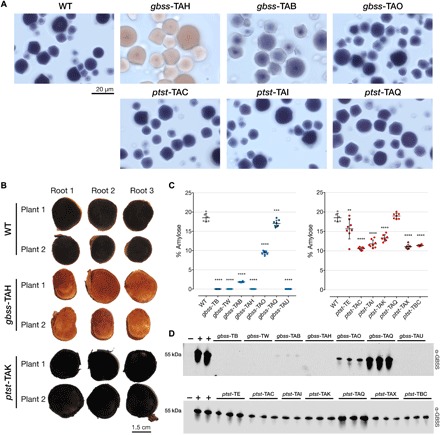
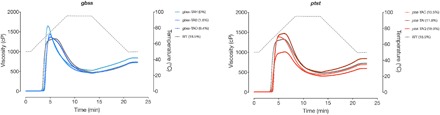
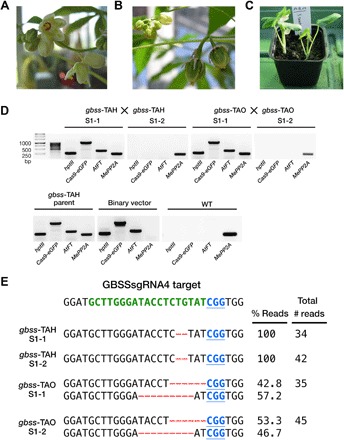
Crop diversification required to meet demands for food security and industrial use is often challenged by breeding time and amenability of varieties to genome modification. Cassava is one such crop. Grown for its large starch-rich storage roots, it serves as a staple food and a commodity in the multibillion-dollar starch industry. Starch is composed of the glucose polymers amylopectin and amylose, with the latter strongly influencing the physicochemical properties of starch during cooking and processing. We demonstrate that CRISPR-Cas9 (clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9)-mediated targeted mutagenesis of two genes involved in amylose biosynthesis, PROTEIN TARGETING TO STARCH (PTST1) or GRANULE BOUND STARCH SYNTHASE (GBSS), can reduce or eliminate amylose content in root starch. Integration of the Arabidopsis FLOWERING LOCUS T gene in the genome-editing cassette allowed us to accelerate flowering-an event seldom seen under glasshouse conditions. Germinated seeds yielded S1, a transgene-free progeny that inherited edited genes. This attractive new plant breeding technique for modified cassava could be extended to other crops to provide a suite of novel varieties with useful traits for food and industrial applications.
Figures
References
-
- H. Ceballos, C. H. Hershey, Cassava (Manihot esculenta Crantz), in Genetic Improvement of Tropical Crops, H. Campos, P. D. S. Caligari, Eds. (Springer International Publishing AG, 2017), pp. 129–180.
-
- R. Howeler, N. Lutaladio, G. Thomas, Save and Grow: Cassava. A Guide to Sustainable Production Intensification (FAO, 2013).
-
- Jansson C., Westerbergh A., Zhang J., Hu X., Sun C., Cassava, a potential biofuel crop in (the) People’s Republic of China. Appl. Energy 86, S95–S99 (2009).
-
- Østerberg J. T., Xiang W., Olsen L. I., Edenbrandt A. K., Vedel S. E., Christiansen A., Landes X., Andersen M. M., Pagh P., Sandøe P., Nielsen J., Christensen S. B., Thorsen B. J., Kappel K., Gamborg C., Palmgren M., Accelerating the domestication of new crops: Feasibility and approaches. Trends Plant Sci. 22, 373–384 (2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
