

Oxidative Damage Control during Decay of Wood by Brown Rot Fungus Using Oxygen Radicals
- PMID: 30194102
- PMCID: PMC6210117
- DOI: 10.1128/AEM.01937-18
Oxidative Damage Control during Decay of Wood by Brown Rot Fungus Using Oxygen Radicals
Abstract
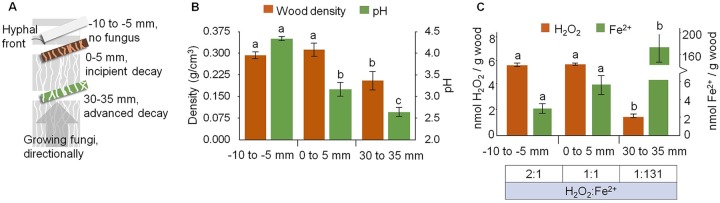
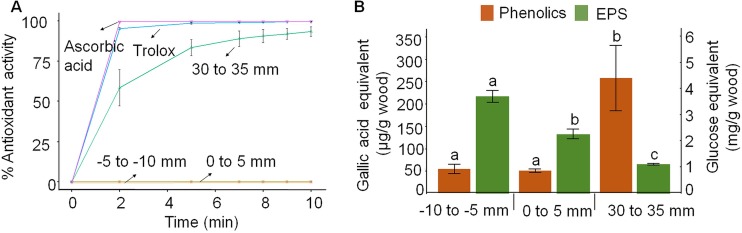
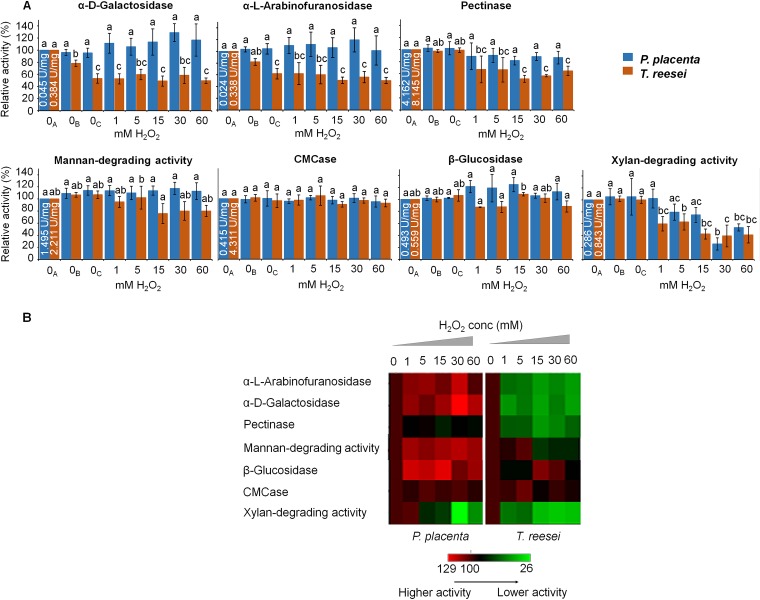
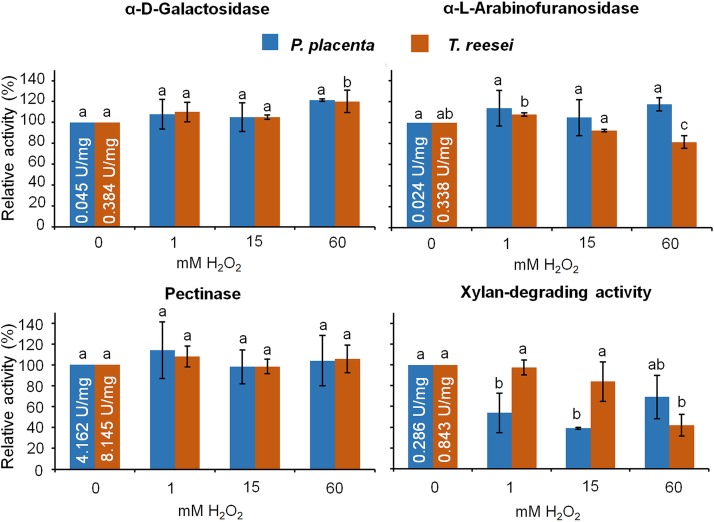
Brown rot wood-degrading fungi deploy reactive oxygen species (ROS) to loosen plant cell walls and enable selective polysaccharide extraction. These ROS, including Fenton-generated hydroxyl radicals (HO˙), react with little specificity and risk damaging hyphae and secreted enzymes. Recently, it was shown that brown rot fungi reduce this risk, in part, by differentially expressing genes involved in HO˙ generation ahead of those coding carbohydrate-active enzymes (CAZYs). However, there are notable exceptions to this pattern, and we hypothesized that brown rot fungi would require additional extracellular mechanisms to limit ROS damage. To assess this, we grew Postia placenta directionally on wood wafers to spatially segregate early from later decay stages. Extracellular HO˙ production (avoidance) and quenching (suppression) capacities among the stages were analyzed, along with the ability of secreted CAZYs to maintain activity postoxidation (tolerance). First, we found that H2O2 and Fe2+ concentrations in the extracellular environment were conducive to HO˙ production in early (H2O2:Fe2+ ratio 2:1) but not later (ratio 1:131) stages of decay. Second, we found that ABTS radical cation quenching (antioxidant capacity) was higher in later decay stages, coincident with higher fungal phenolic concentrations. Third, by surveying enzyme activities before/after exposure to Fenton-generated HO˙, we found that CAZYs secreted early, amid HO˙, were more tolerant of oxidative stress than those expressed later and were more tolerant than homologs in the model CAZY producer Trichoderma reesei Collectively, this indicates that P. placenta uses avoidance, suppression, and tolerance mechanisms, extracellularly, to complement intracellular differential expression, enabling this brown rot fungus to use ROS to degrade wood.IMPORTANCE Wood is one of the largest pools of carbon on Earth, and its decomposition is dominated in most systems by fungi. Wood-degrading fungi specialize in extracting sugars bound within lignin, either by removing lignin first (white rot) or by using Fenton-generated reactive oxygen species (ROS) to "loosen" wood cell walls, enabling selective sugar extraction (brown rot). Although white rot lignin-degrading pathways are well characterized, there are many uncertainties in brown rot fungal mechanisms. Our study addressed a key uncertainty in how brown rot fungi deploy ROS without damaging themselves or the enzymes they secrete. In addition to revealing differentially expressed genes to promote ROS generation only in early decay, our study revealed three spatial control mechanisms to avoid/tolerate ROS: (i) constraining Fenton reactant concentrations (H2O2, Fe2+), (ii) quenching ROS via antioxidants, and (iii) secreting ROS-tolerant enzymes. These results not only offer insight into natural decomposition pathways but also generate targets for biotechnological development.
Keywords: Fenton reaction; antioxidant capacity; glycosyl hydrolases; hydroxyl radicals; oxidative stress tolerance; wood decay.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Gilbertson RL, Ryvarden L. 1986. North American polypores. Fungiflora, Oslo, Norway.
-
- Zabel RA, Morrell JJ. 1992. Wood microbiology: decay and its prevention. Academic Press, San Diego, CA.
-
- Eriksson KE, Blanchette RA, Ander P. 1990. Microbial and enzymatic degradation of wood and wood components. Springer, Berlin, Germany.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
