

RNA Polymerase II Activity of Type 3 Pol III Promoters
- PMID: 30195753
- PMCID: PMC6023835
- DOI: 10.1016/j.omtn.2018.05.001
RNA Polymerase II Activity of Type 3 Pol III Promoters
Abstract
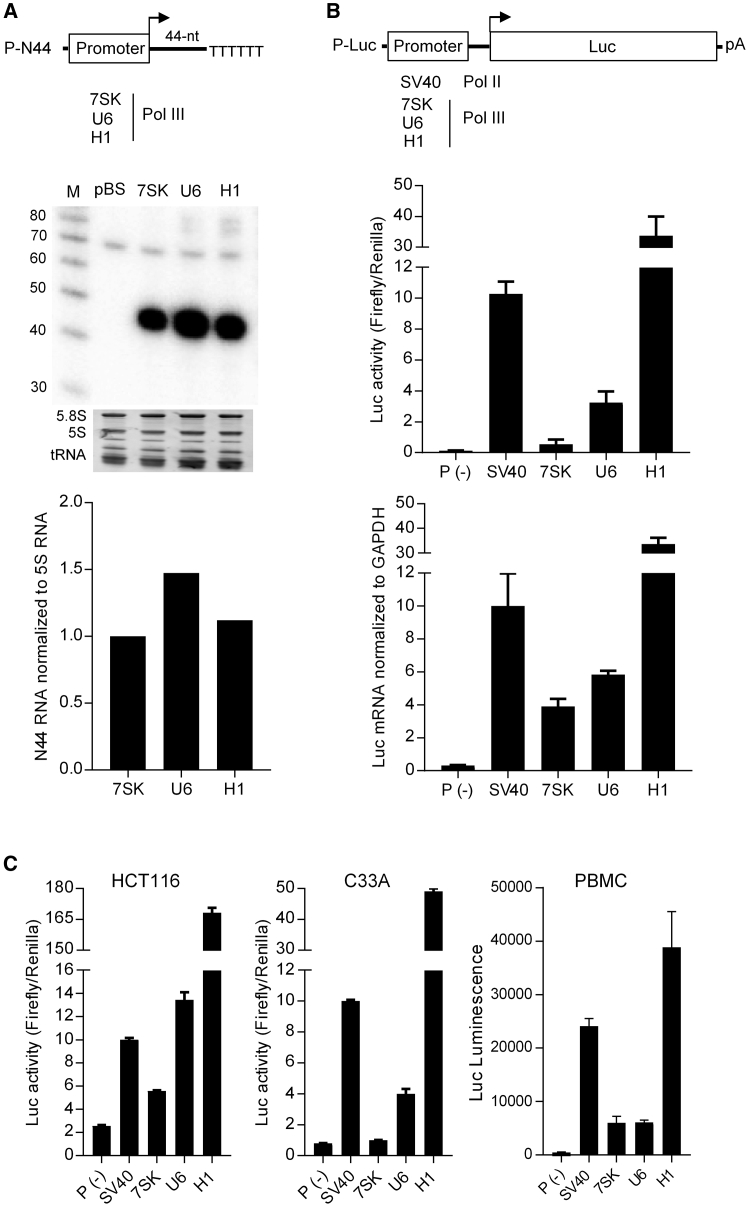
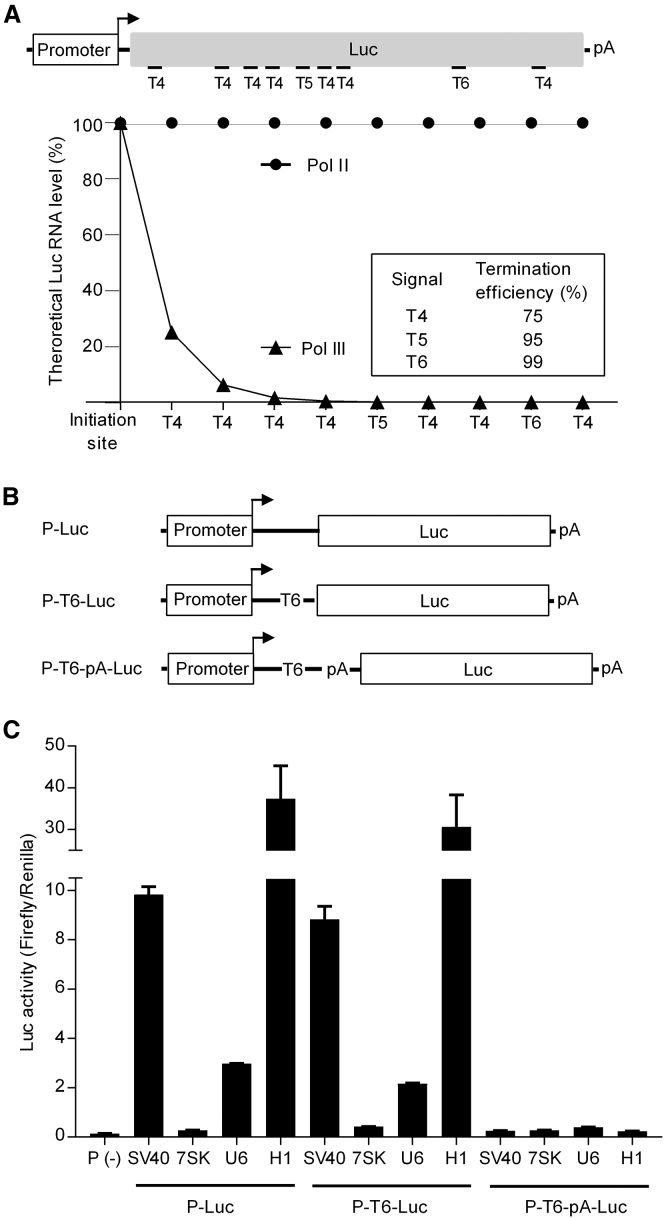
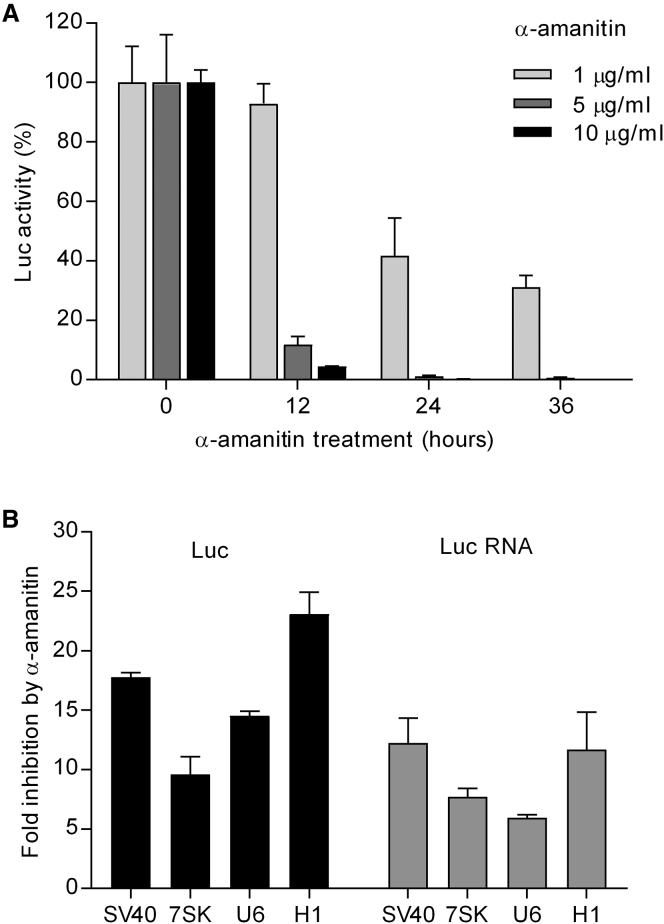
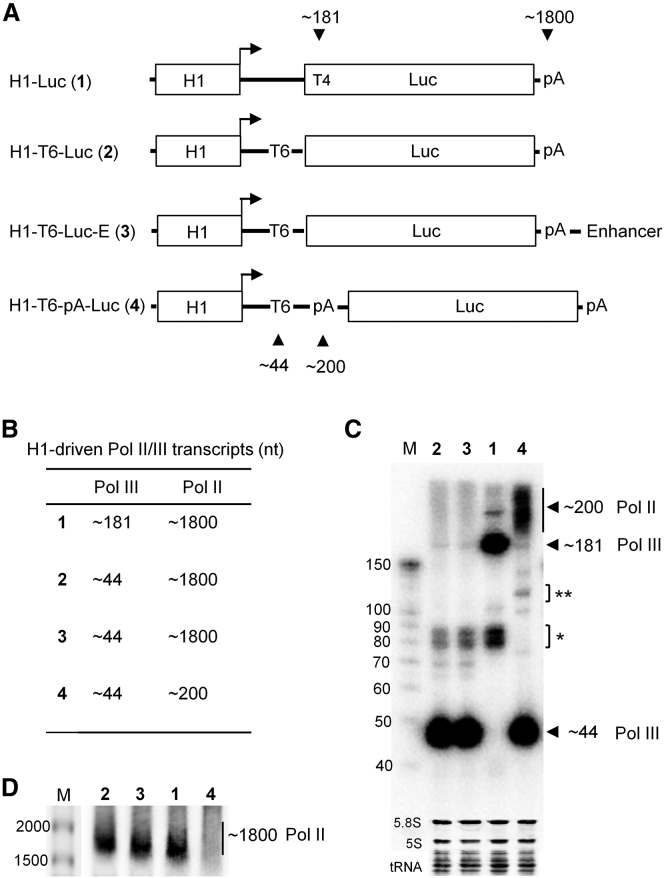
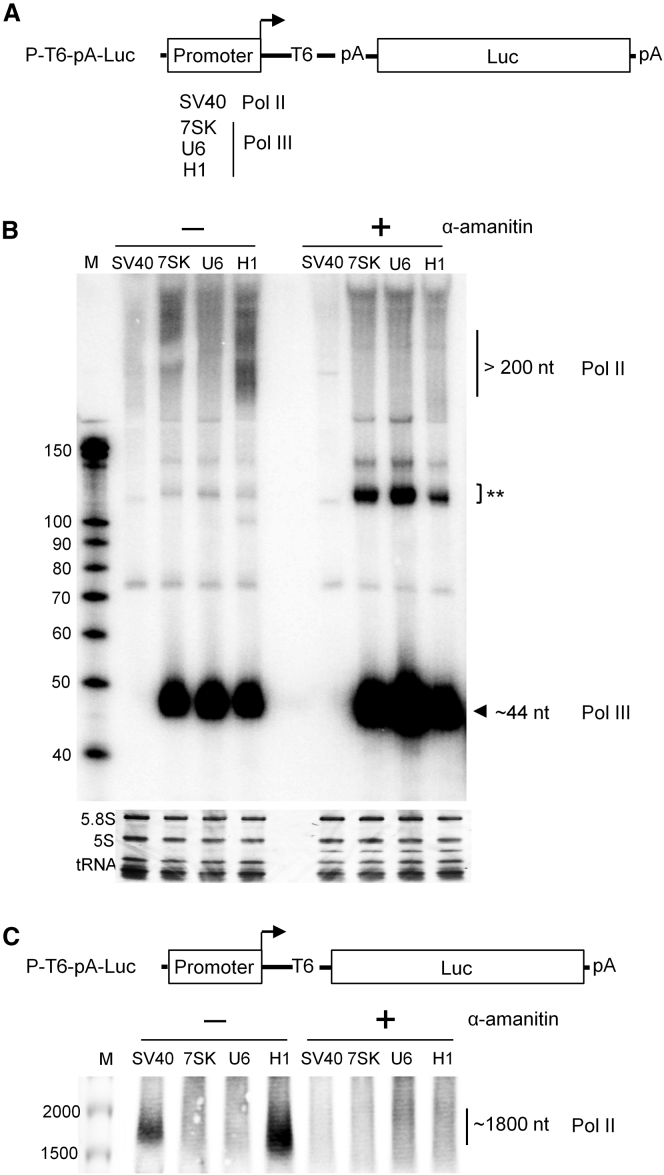
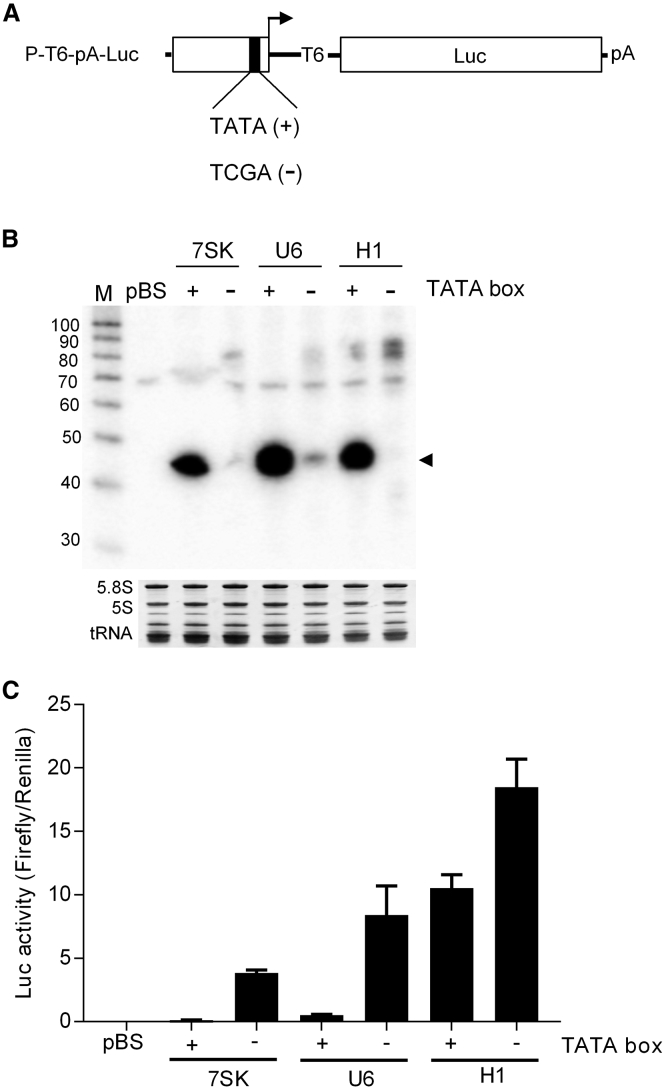
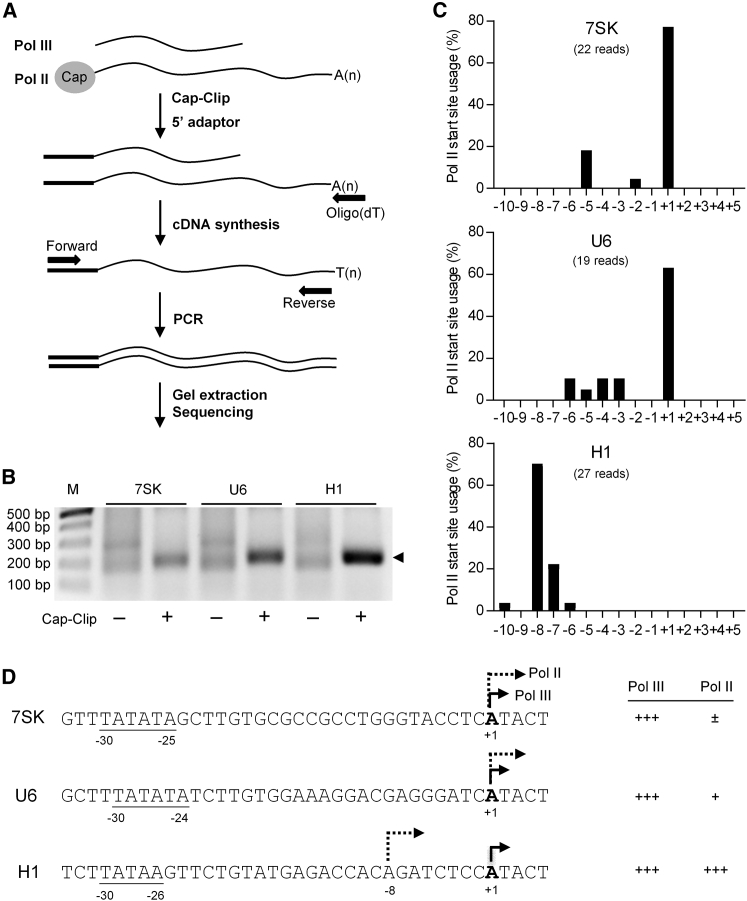
In eukaryotes, three RNA polymerases (Pol I, II, and III) are responsible for the transcription of distinct subsets of genes. Gene-external type 3 Pol III promoters use defined transcription start and termination sites, and they are, therefore, widely used for small RNA expression, including short hairpin RNAs in RNAi applications and guide RNAs in CRISPR-Cas systems. We report that all three commonly used human Pol III promoters (7SK, U6, and H1) mediate luciferase reporter gene expression, which indicates Pol II activity, but to a different extent (H1 ≫ U6 > 7SK). We demonstrate that these promoters can recruit Pol II for transcribing extended messenger transcripts. Intriguingly, selective inhibition of Pol II stimulates the Pol III activity and vice versa, suggesting that two polymerase complexes compete for promoter usage. Pol II initiates transcription at the regular Pol III start site on the 7SK and U6 promoters, but Pol II transcription on the most active H1 promoter starts 8 nt upstream of the Pol III start site. This study provides functional evidence for the close relationship of Pol II and Pol III transcription. These mechanistic insights are important for optimal use of Pol III promoters, and they offer additional flexibility for biotechnology applications of these genetic elements.
Keywords: Pol II; Pol III; dual-polymerase activity; promoter competition; type 3 Pol III promoters.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Vannini A., Cramer P. Conservation between the RNA polymerase I, II, and III transcription initiation machineries. Mol. Cell. 2012;45:439–446. - PubMed
-
- Cramer P., Armache K.J., Baumli S., Benkert S., Brueckner F., Buchen C., Damsma G.E., Dengl S., Geiger S.R., Jasiak A.J. Structure of eukaryotic RNA polymerases. Annu. Rev. Biophys. 2008;37:337–352. - PubMed
-
- Oler A.J., Alla R.K., Roberts D.N., Wong A., Hollenhorst P.C., Chandler K.J., Cassiday P.A., Nelson C.A., Hagedorn C.H., Graves B.J., Cairns B.R. Human RNA polymerase III transcriptomes and relationships to Pol II promoter chromatin and enhancer-binding factors. Nat. Struct. Mol. Biol. 2010;17:620–628. - PMC - PubMed
-
- Schramm L., Hernandez N. Recruitment of RNA polymerase III to its target promoters. Genes Dev. 2002;16:2593–2620. - PubMed
-
- Murphy S., Di Liegro C., Melli M. The in vitro transcription of the 7SK RNA gene by RNA polymerase III is dependent only on the presence of an upstream promoter. Cell. 1987;51:81–87. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
