

Trypanosomatid Deoxyhypusine Synthase Activity Is Dependent on Shared Active-Site Complementation between Pseudoenzyme Paralogs
- PMID: 30197036
- PMCID: PMC6221947
- DOI: 10.1016/j.str.2018.07.012
Trypanosomatid Deoxyhypusine Synthase Activity Is Dependent on Shared Active-Site Complementation between Pseudoenzyme Paralogs
Abstract
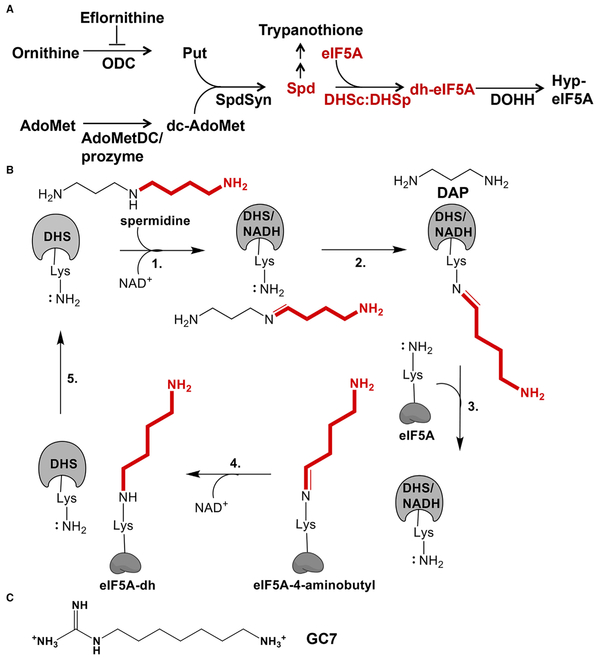
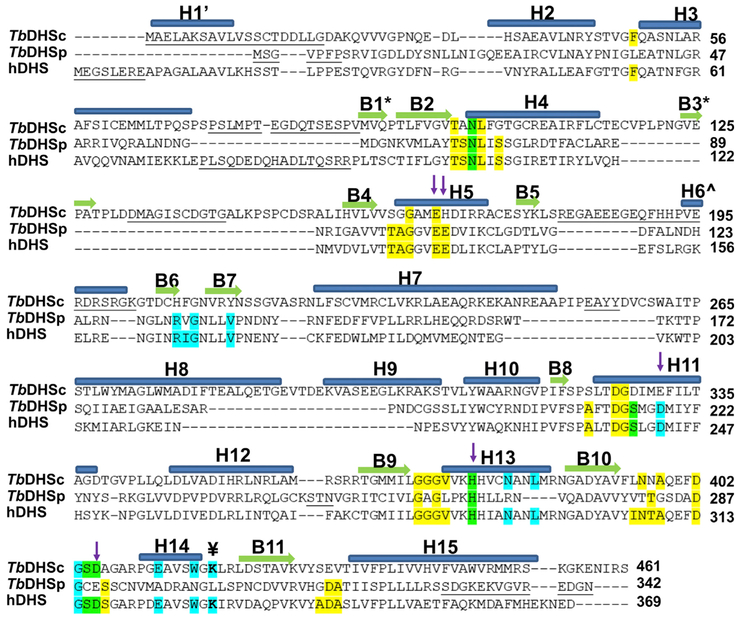
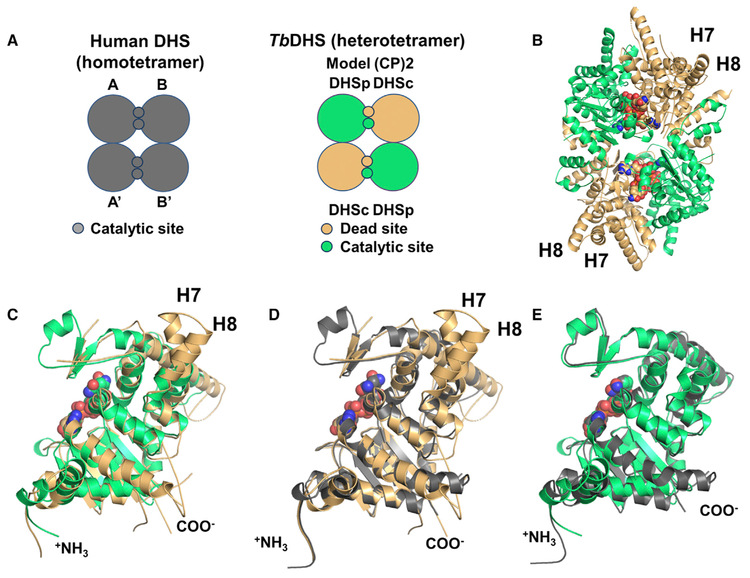
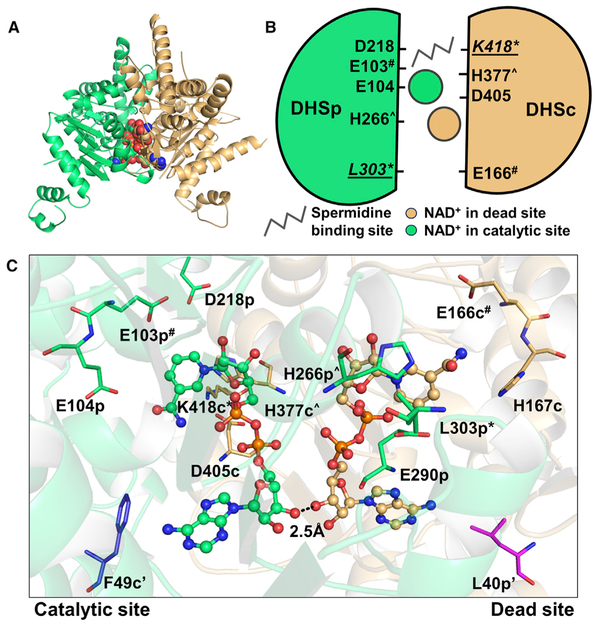
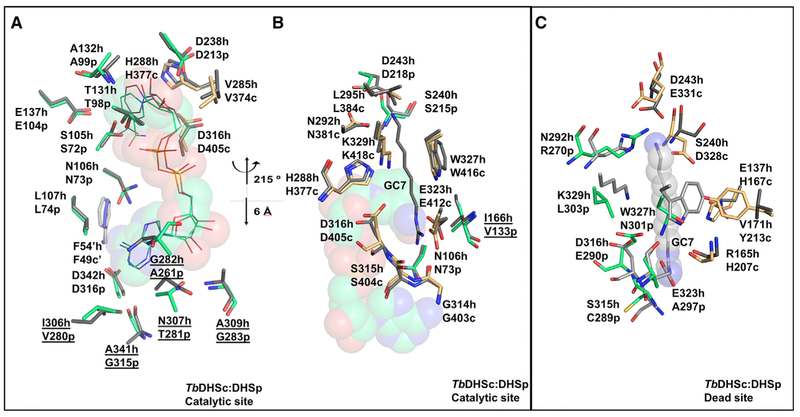
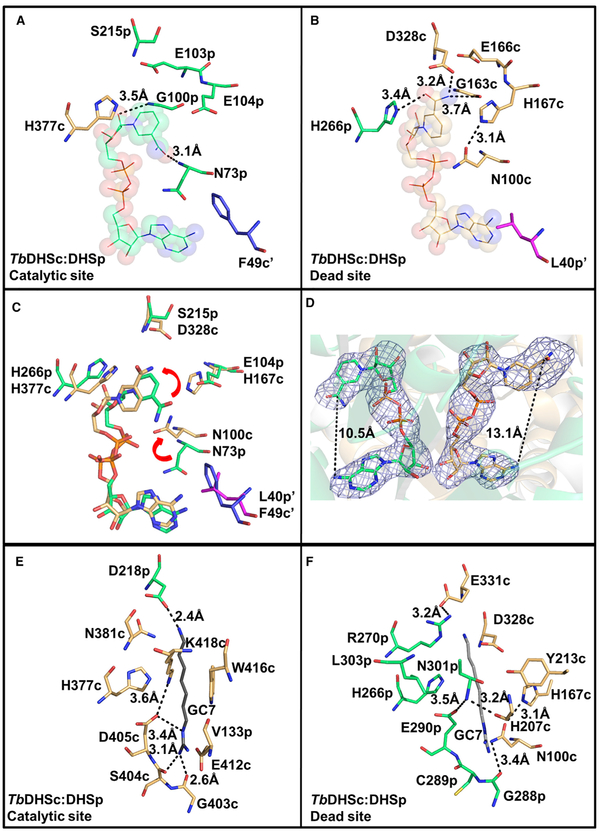
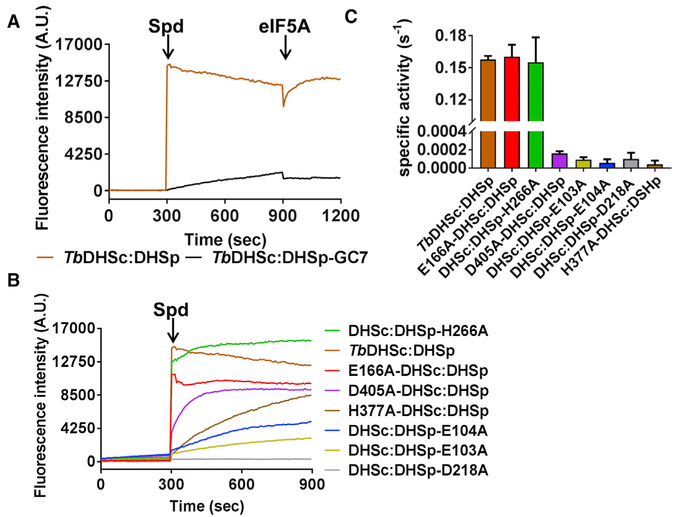
Trypanosoma brucei is a neglected tropical disease endemic to Africa. The polyamine spermidine is essential for post-translational hypusine modification of eukaryotic initiation factor 5A (eIF5A), which is catalyzed by deoxyhypusine synthase (TbDHS). In trypanosomatids, deoxyhypusine synthase (DHS) activity is dependent on heterotetramer formation between two paralogs, DHSc and DHSp, both with minimal activity on their own due to missing catalytic residues. We determined the X-ray structure of TbDHS showing a single functional shared active site is formed at the DHSc/DHSp heterodimer interface, with deficiencies in one subunit complemented by the other. Each heterodimer contains two NAD+ binding sites, one housed in the functional catalytic site and the second bound in a remnant dead site that lacks key catalytic residues. Functional analysis of these sites by site-directed mutagenesis identified long-range contributions to the catalytic site from the dead site. Differences between trypanosomatid and human DHS that could be exploited for drug discovery were identified.
Keywords: Trypanosoma brucei; deoxyhypusine synthase; eIF5A; hypusine; polyamines; pseudoenzyme; trypanosomatids.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Unique Characteristics of the Parasite Polyamine Pathway.Structure. 2018 Nov 6;26(11):1427-1429. doi: 10.1016/j.str.2018.10.018. Structure. 2018. PMID: 30403992
References
-
- Borek D, Minor W, and Otwinowski Z (2003). Measurement errors and their consequences in protein crystallography. Acta Crystallogr. D Biol. Crystallogr 59,2031–2038. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
