

Human Dystrophin Structural Changes upon Binding to Anionic Membrane Lipids
- PMID: 30197181
- PMCID: PMC6170597
- DOI: 10.1016/j.bpj.2018.07.039
Human Dystrophin Structural Changes upon Binding to Anionic Membrane Lipids
Abstract
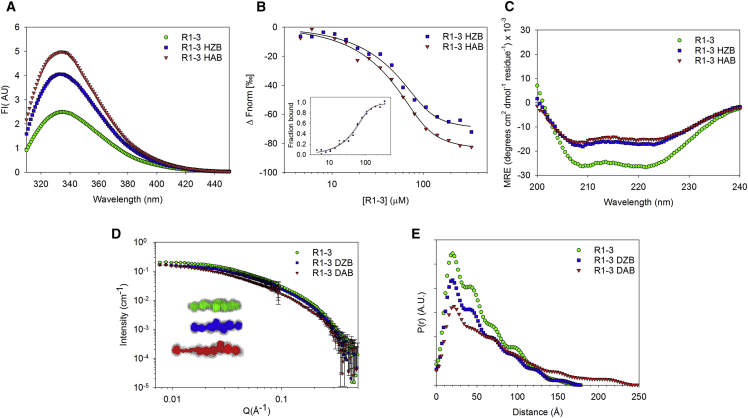
Scaffolding proteins play important roles in supporting the plasma membrane (sarcolemma) of muscle cells. Among them, dystrophin strengthens the sarcolemma through protein-lipid interactions, and its absence due to gene mutations leads to the severe Duchenne muscular dystrophy. Most of the dystrophin protein consists of a central domain made of 24 spectrin-like coiled-coil repeats (R). Using small angle neutron scattering (SANS) and the contrast variation technique, we specifically probed the structure of the three first consecutive repeats 1-3 (R1-3), a part of dystrophin known to physiologically interact with membrane lipids. R1-3 free in solution was compared to its structure adopted in the presence of phospholipid-based bicelles. SANS data for the protein/lipid complexes were obtained with contrast-matched bicelles under various phospholipid compositions to probe the role of electrostatic interactions. When bound to anionic bicelles, large modifications of the protein three-dimensional structure were detected, as revealed by a significant increase of the protein gyration radius from 42 ± 1 to 60 ± 4 Å. R1-3/anionic bicelle complexes were further analyzed by coarse-grained molecular dynamics simulations. From these studies, we report an all-atom model of R1-3 that highlights the opening of the R1 coiled-coil repeat when bound to the membrane lipids. This model is totally in agreement with SANS and click chemistry/mass spectrometry data. We conclude that the sarcolemma membrane anchoring that occurs during the contraction/elongation process of muscles could be ensured by this coiled-coil opening. Therefore, understanding these structural changes may help in the design of rationalized shortened dystrophins for gene therapy. Finally, our strategy opens up new possibilities for structure determination of peripheral and integral membrane proteins not compatible with different high-resolution structural methods.
Copyright © 2018 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures


References
-
- Seddon A.M., Curnow P., Booth P.J. Membrane proteins, lipids and detergents: not just a soap opera. Biochim. Biophys. Acta. 2004;1666:105–117. - PubMed
-
- Petoukhov M.V., Svergun D.I. Analysis of X-ray and neutron scattering from biomacromolecular solutions. Curr. Opin. Struct. Biol. 2007;17:562–571. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
