

Regulation of energy metabolism during early mammalian development: TEAD4 controls mitochondrial transcription
- PMID: 30201685
- PMCID: PMC6198476
- DOI: 10.1242/dev.162644
Regulation of energy metabolism during early mammalian development: TEAD4 controls mitochondrial transcription
Abstract
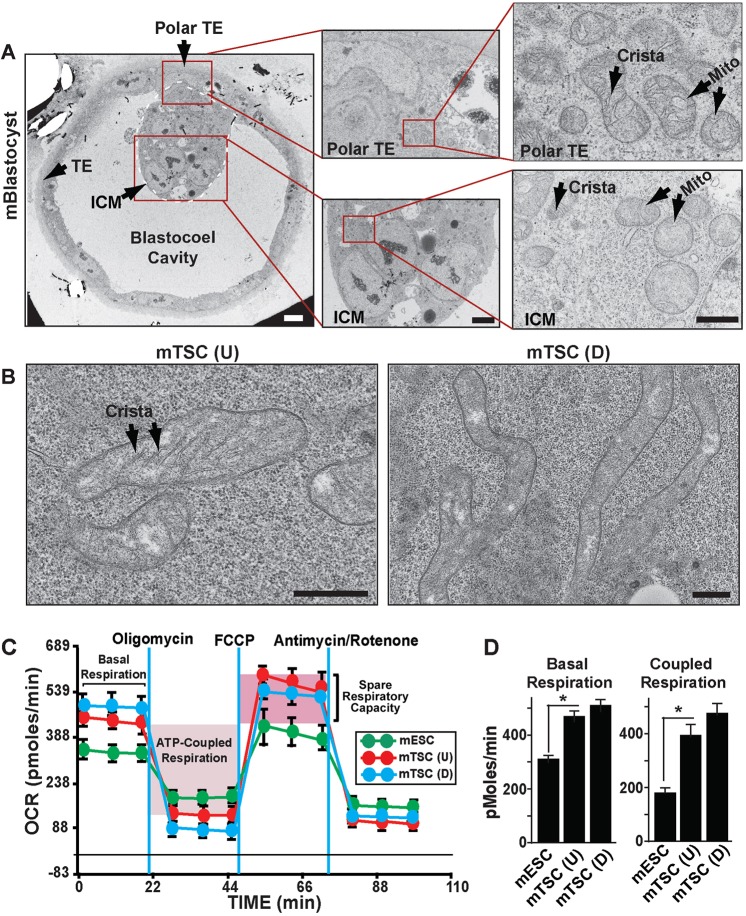
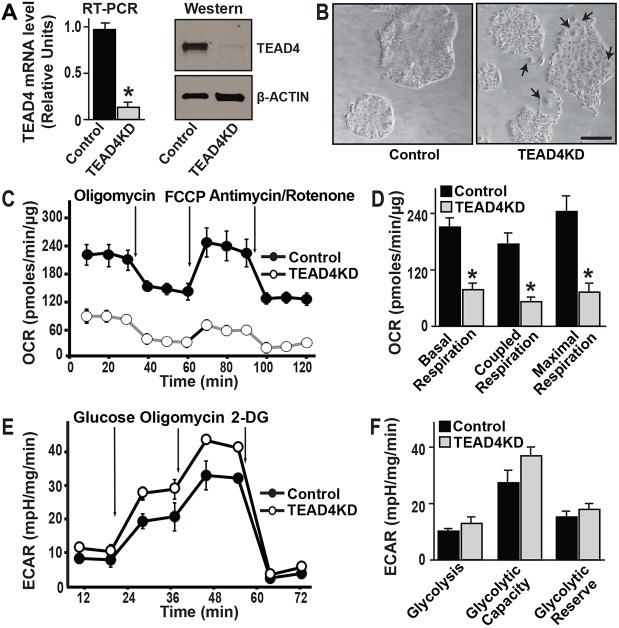
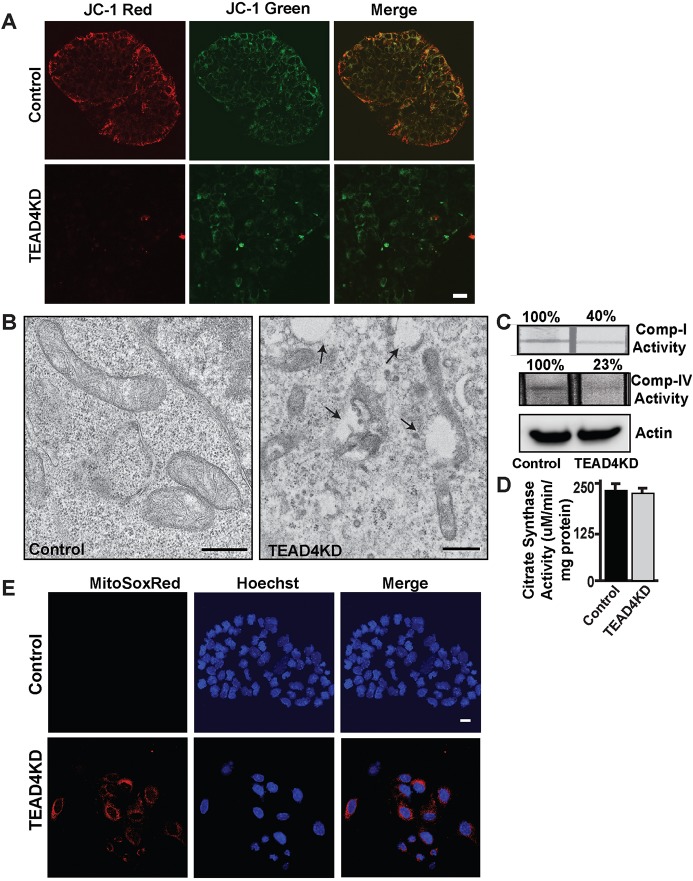
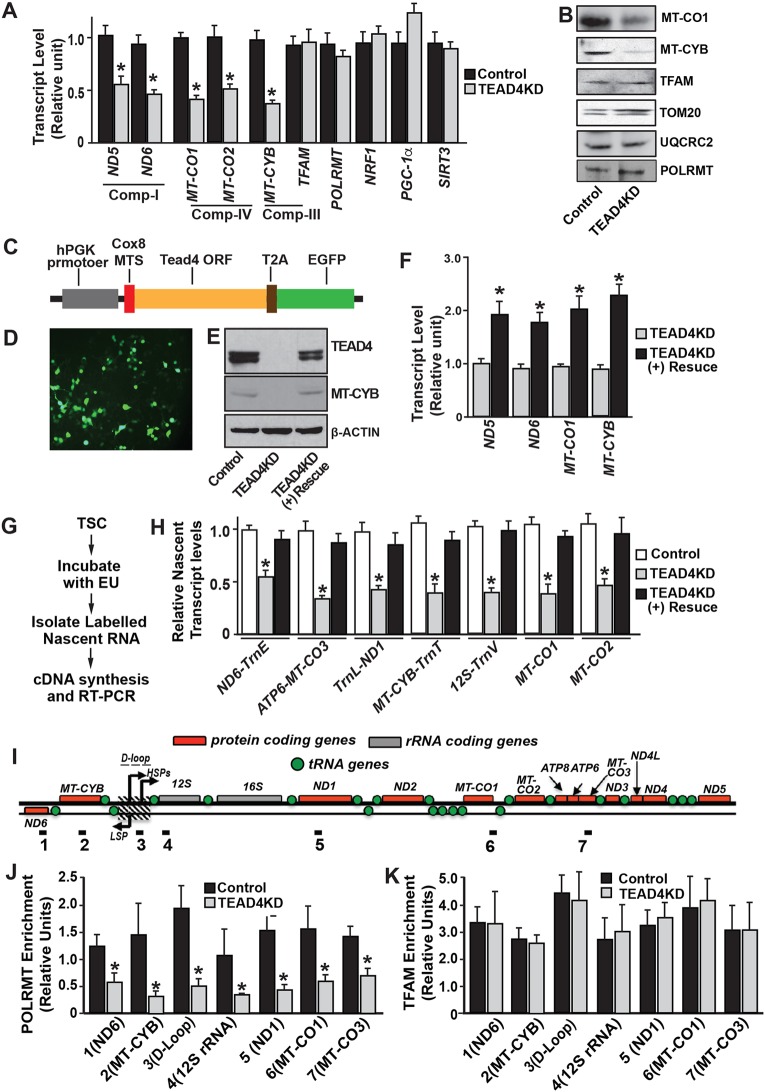
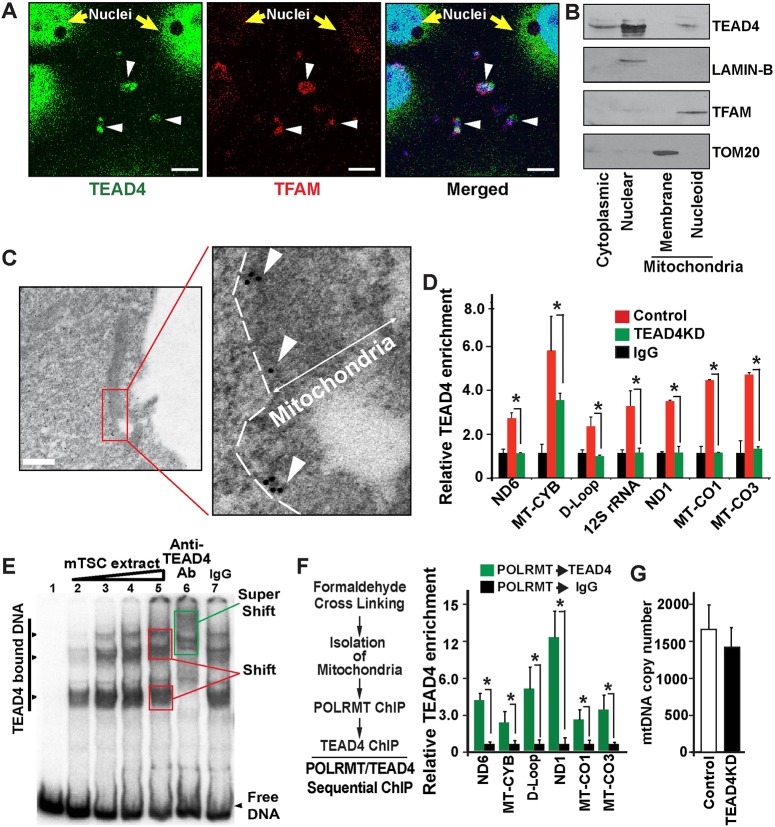
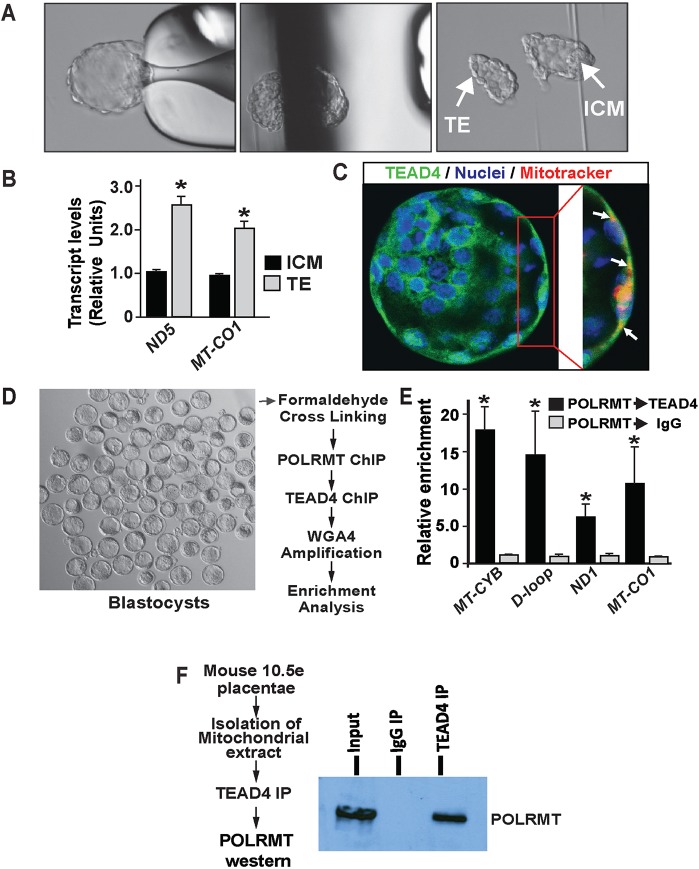
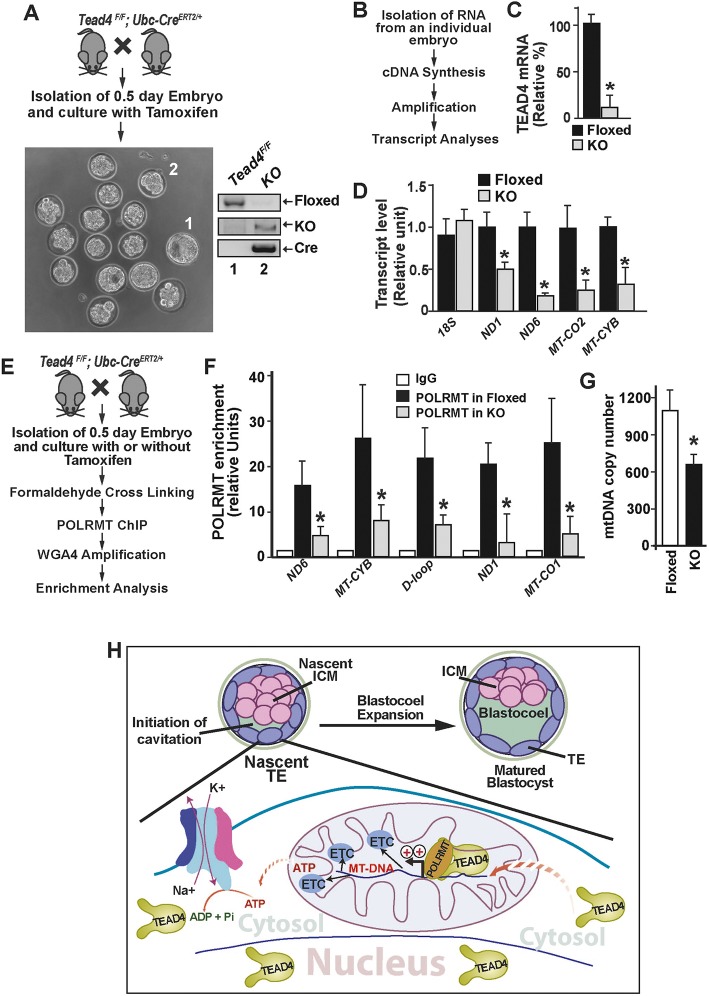
Early mammalian development is crucially dependent on the establishment of oxidative energy metabolism within the trophectoderm (TE) lineage. Unlike the inner cell mass, TE cells enhance ATP production via mitochondrial oxidative phosphorylation (OXPHOS) and this metabolic preference is essential for blastocyst maturation. However, molecular mechanisms that regulate establishment of oxidative energy metabolism in TE cells are incompletely understood. Here, we show that conserved transcription factor TEAD4, which is essential for pre-implantation mammalian development, regulates this process by promoting mitochondrial transcription. In developing mouse TE and TE-derived trophoblast stem cells (TSCs), TEAD4 localizes to mitochondria, binds to mitochondrial DNA (mtDNA) and facilitates its transcription by recruiting mitochondrial RNA polymerase (POLRMT). Loss of TEAD4 impairs recruitment of POLRMT, resulting in reduced expression of mtDNA-encoded electron transport chain components, thereby inhibiting oxidative energy metabolism. Our studies identify a novel TEAD4-dependent molecular mechanism that regulates energy metabolism in the TE lineage to ensure mammalian development.
Keywords: Electron Transport Chain; Mammalian development; Mitochondrial transcription; POLRMT; TEAD4; Trophoblast stem cell.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
Tead4 is required for specification of trophectoderm in pre-implantation mouse embryos.Mech Dev. 2008 Mar-Apr;125(3-4):270-83. doi: 10.1016/j.mod.2007.11.002. Epub 2007 Nov 17. Mech Dev. 2008. PMID: 18083014
-
Altered subcellular localization of transcription factor TEAD4 regulates first mammalian cell lineage commitment.Proc Natl Acad Sci U S A. 2012 May 8;109(19):7362-7. doi: 10.1073/pnas.1201595109. Epub 2012 Apr 23. Proc Natl Acad Sci U S A. 2012. PMID: 22529382 Free PMC article.
-
TEAD4 ensures postimplantation development by promoting trophoblast self-renewal: An implication in early human pregnancy loss.Proc Natl Acad Sci U S A. 2020 Jul 28;117(30):17864-17875. doi: 10.1073/pnas.2002449117. Epub 2020 Jul 15. Proc Natl Acad Sci U S A. 2020. PMID: 32669432 Free PMC article.
-
Mechanisms of trophectoderm fate specification in preimplantation mouse development.Dev Growth Differ. 2010 Apr;52(3):263-73. doi: 10.1111/j.1440-169X.2009.01158.x. Epub 2010 Jan 20. Dev Growth Differ. 2010. PMID: 20100249 Review.
-
The transcription machinery in mammalian mitochondria.Biochim Biophys Acta. 2004 Dec 6;1659(2-3):148-52. doi: 10.1016/j.bbabio.2004.10.003. Biochim Biophys Acta. 2004. PMID: 15576046 Review.
Cited by
-
Arginine: at the crossroads of nitrogen metabolism.EMBO J. 2025 Mar;44(5):1275-1293. doi: 10.1038/s44318-025-00379-3. Epub 2025 Feb 7. EMBO J. 2025. PMID: 39920310 Free PMC article. Review.
-
Mitoguardin 1 and 2 promote granulosa cell proliferation by activating AKT and regulating the Hippo-YAP1 signaling pathway.Cell Death Dis. 2023 Nov 27;14(11):779. doi: 10.1038/s41419-023-06312-y. Cell Death Dis. 2023. PMID: 38012141 Free PMC article.
-
Editorial: The proceedings of mitochondria apoptosis and cancer (MAC 2021) virtual symposium.Front Cell Dev Biol. 2023 Jan 4;10:1118314. doi: 10.3389/fcell.2022.1118314. eCollection 2022. Front Cell Dev Biol. 2023. PMID: 36684418 Free PMC article. No abstract available.
-
Embryo and Its Mitochondria.Antioxidants (Basel). 2021 Jan 20;10(2):139. doi: 10.3390/antiox10020139. Antioxidants (Basel). 2021. PMID: 33498182 Free PMC article. Review.
-
OCT4 induces embryonic pluripotency via STAT3 signaling and metabolic mechanisms.Proc Natl Acad Sci U S A. 2021 Jan 19;118(3):e2008890118. doi: 10.1073/pnas.2008890118. Proc Natl Acad Sci U S A. 2021. PMID: 33452132 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
