

Tissue signals imprint ILC2 identity with anticipatory function
- PMID: 30201992
- PMCID: PMC6202223
- DOI: 10.1038/s41590-018-0201-4
Tissue signals imprint ILC2 identity with anticipatory function
Abstract
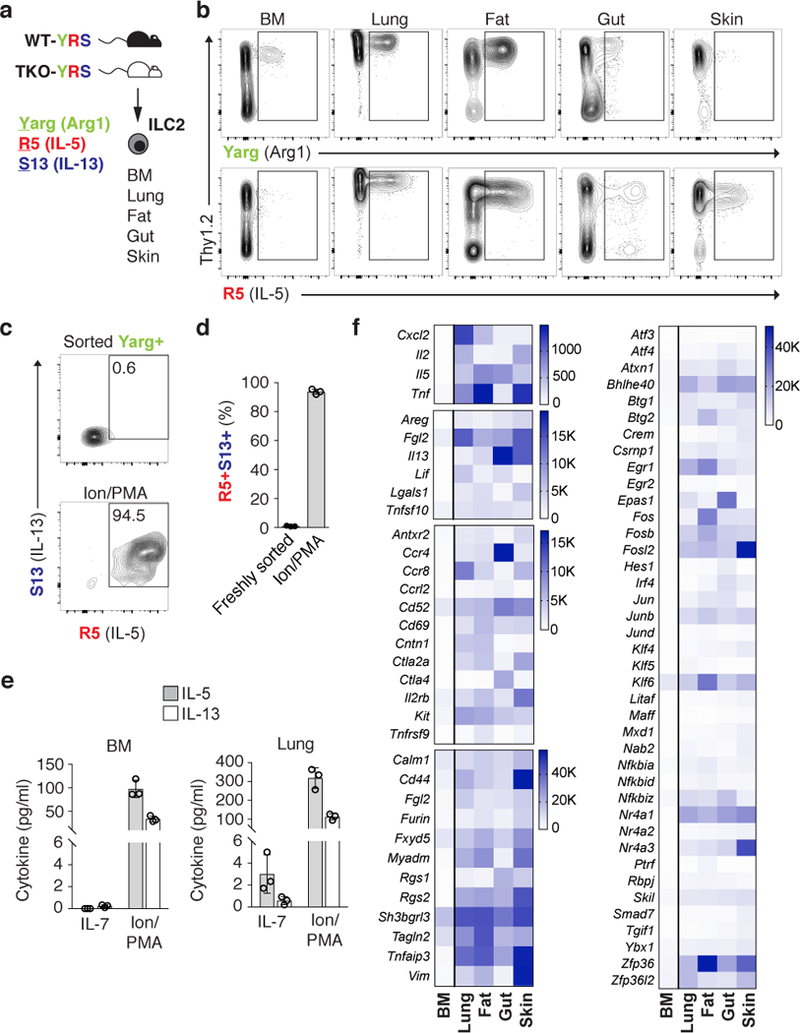
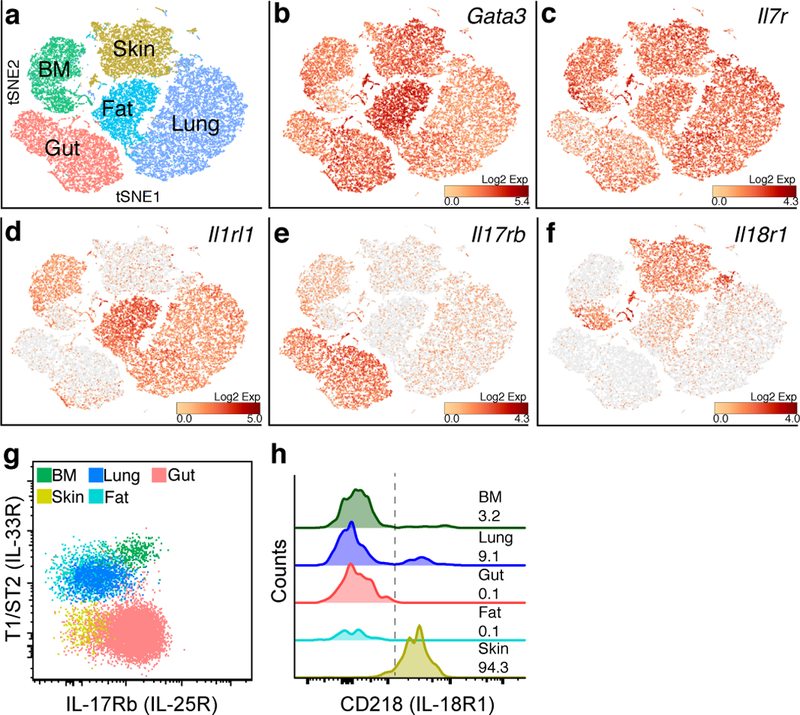
Group 2 innate lymphoid cells (ILC2s) are distributed systemically and produce type 2 cytokines in response to a variety of stimuli, including the epithelial cytokines interleukin (IL)-25, IL-33, and thymic stromal lymphopoietin (TSLP). Transcriptional profiling of ILC2s from different tissues, however, grouped ILC2s according to their tissue of origin, even in the setting of combined IL-25-, IL-33-receptor-, and TSLP-receptor-deficiency. Single-cell profiling confirmed a tissue-organizing transcriptome and identified ILC2 subsets expressing distinct activating receptors, including the major subset of skin ILC2s, which were activated preferentially by IL-18. Tissue ILC2 subsets were unaltered in number and expression in germ-free mice, suggesting that endogenous, tissue-derived signals drive the maturation of ILC2 subsets by controlling expression of distinct patterns of activating receptors, thus anticipating tissue-specific perturbations occurring later in life.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Competing interests: None declared.
Figures





Comment in
-
Mysterious ILC2 tissue adaptation.Nat Immunol. 2018 Oct;19(10):1042-1044. doi: 10.1038/s41590-018-0214-z. Nat Immunol. 2018. PMID: 30201993 No abstract available.
References
-
- Klose CSN & Artis D Innate lymphoid cells as regulators of immunity, inflammation and tissue homeostasis. Nat. Immunol 17, 765–774 (2016). - PubMed
-
- Klein Wolterink RGJ et al. Pulmonary innate lymphoid cells are major producers of IL-5 and IL-13 in murine models of allergic asthma. Eur. J. Immunol 42, 1106–1116 (2012). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
