

Cell-wall-degrading enzymes produced in vitro and in vivo by Rhizoctonia solani, the causative fungus of peanut sheath blight
- PMID: 30202660
- PMCID: PMC6129149
- DOI: 10.7717/peerj.5580
Cell-wall-degrading enzymes produced in vitro and in vivo by Rhizoctonia solani, the causative fungus of peanut sheath blight
Abstract
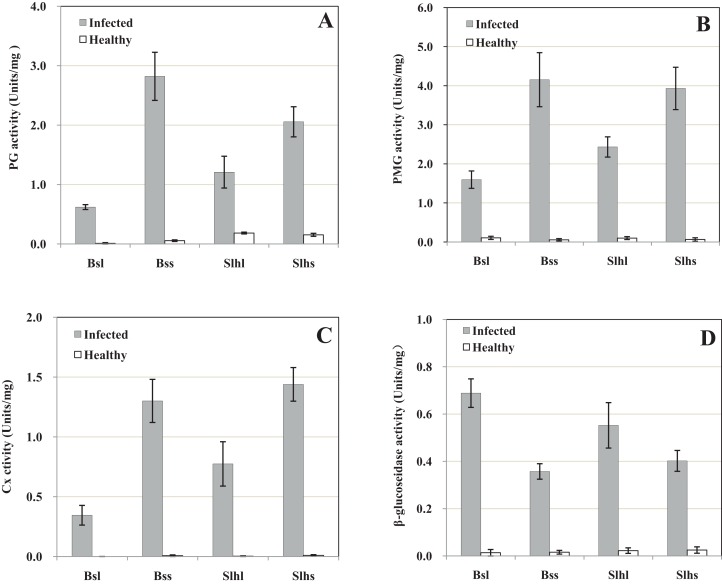
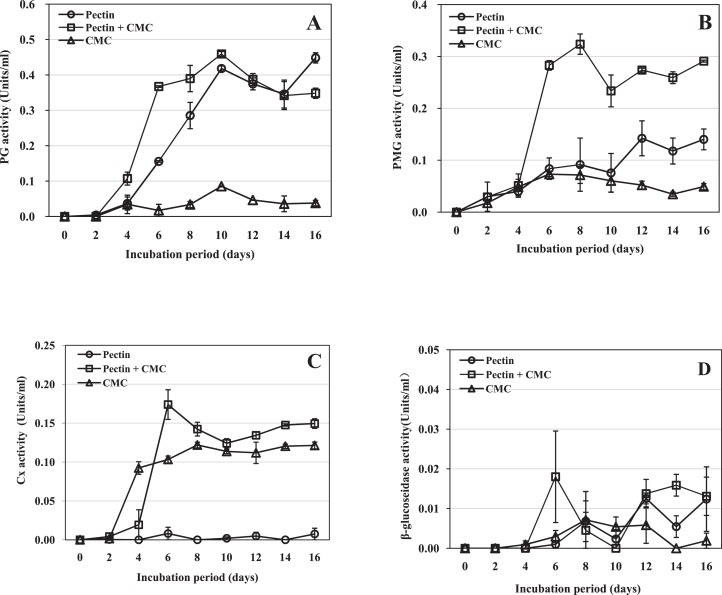
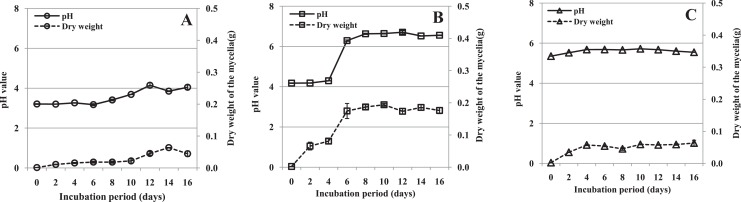
Rhizoctonia solani causes the disease peanut sheath blight, involving symptoms of maceration and necrosis of infected tissue, mainly caused by cell-wall-degrading enzymes (CWDEs). This study investigated the production of CWDEs including polygalacturonase (PG), polymethyl-galacturonase (PMG), cellulase (Cx) and β-glucosidase by R. solani in vitro (in liquid culture) and in vivo (in peanut plants). Significant PG, PMG, Cx and β-glucosidase activities were detected in infected tissues including stalk and leaves of Baisha and Silihong peanut cultivars. Extracts of healthy tissue showed little or no such activities. In shaken liquid cultures of R. solani in medium containing pectin or pectin plus carboxymethyl cellulose (CMC) as the carbon source(s), PG and PMG were notably active. Significant Cx activity was detected in cultures with CMC or pectin plus CMC as the carbon source(s). However, only a very low level of β-glucosidase activity was observed in cultures with any of the tested carbon sources. An increase of pH was recorded in decayed peanut tissues and liquid culture filtrates; the filtrate pH and fungal growth positively correlated. The fungal growth and/or pH were important factors for the production of PG, PMG and Cx in culture with pectin plus CMC as the carbon source. A single active PG isozyme with isoelectric point around 9.2 was detected in culture filtrates and in infected peanut tissues by the method of isoelectric focusing electrophoresis. The crude enzymes extracted from liquid culture of R. solani induced decay of healthy peanut leaves.
Keywords: Cell-wall-degrading enzymes; IEF; Peanut; Peanut sheath blight disease; Rhizoctonia solani.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Amit KC, Shubha C, Sushmita C, Shridha C. In vitro and in vivo production of polygalacturonase, polymethylgalacturonase and cellulase enzymes by Alternaria solani at different incubation periods. African Journal of Plant Science. 2014;8(5):248–253. doi: 10.5897/ajps2013.1114. - DOI
-
- Anderson JR, Bentley S, Irwin JAG, Mackie JM, Neate S, Pattemore JA. Characterisation of Rhizoctonia solani isolates causing root canker of lucerne in Australia. Australasian Plant Pathology. 2004;33(2):241–247. doi: 10.1071/ap04012. - DOI
-
- Brien PAO, Zamani M. Production of pectic enzymes by barepatch isolates of Rhizoctonia solani AG 8. Australasian Plant Pathology. 2003;32(1):65–72. doi: 10.1071/ap02073. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
