

The Neurocognitive Basis of Spatial Reorientation
- PMID: 30205055
- PMCID: PMC6161705
- DOI: 10.1016/j.cub.2018.04.057
The Neurocognitive Basis of Spatial Reorientation
Abstract
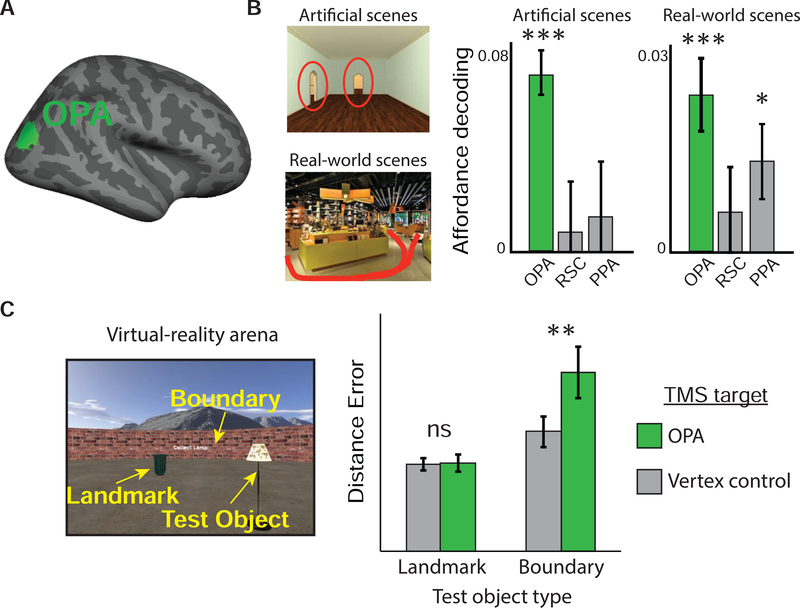
The ability to recover one's bearings when lost is a skill that is fundamental for spatial navigation. We review the cognitive and neural mechanisms that underlie this ability, with the aim of linking together previously disparate findings from animal behavior, human psychology, electrophysiology, and cognitive neuroscience. Behavioral work suggests that reorientation involves two key abilities: first, the recovery of a spatial reference frame (a cognitive map) that is appropriate to the current environment; and second, the determination of one's heading and location relative to that reference frame. Electrophysiological recording studies, primarily in rodents, have revealed potential correlates of these operations in place, grid, border/boundary, and head-direction cells in the hippocampal formation. Cognitive neuroscience studies, primarily in humans, suggest that the perceptual inputs necessary for these operations are processed by neocortical regions such as the retrosplenial complex, occipital place area and parahippocampal place area, with the retrosplenial complex mediating spatial transformations between the local environment and the recovered spatial reference frame, the occipital place area supporting perception of local boundaries, and the parahippocampal place area processing visual information that is essential for identification of the local spatial context. By combining results across these various literatures, we converge on a unified account of reorientation that bridges the cognitive and neural domains.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures





References
-
- O’Keefe J, and Nadel L (1978). The hippocampus as a cognitive map (Clarendon Press Oxford; ).
-
- Gallistel CR (1990). The organization of learning (The MIT Press; ).
-
- Trullier O, Wiener SI, Berthoz A, and Meyer JA (1997). Biologically based artificial navigation systems: Review and prospects. Prog. Neurobiol 51, 483–544. - PubMed
-
- Kuipers B (2000). The spatial semantic hierarchy. Artif. Intell 119, 191–233.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
