

Molecular Dynamics Simulations of Kir2.2 Interactions with an Ensemble of Cholesterol Molecules
- PMID: 30205899
- PMCID: PMC6170799
- DOI: 10.1016/j.bpj.2018.07.041
Molecular Dynamics Simulations of Kir2.2 Interactions with an Ensemble of Cholesterol Molecules
Abstract
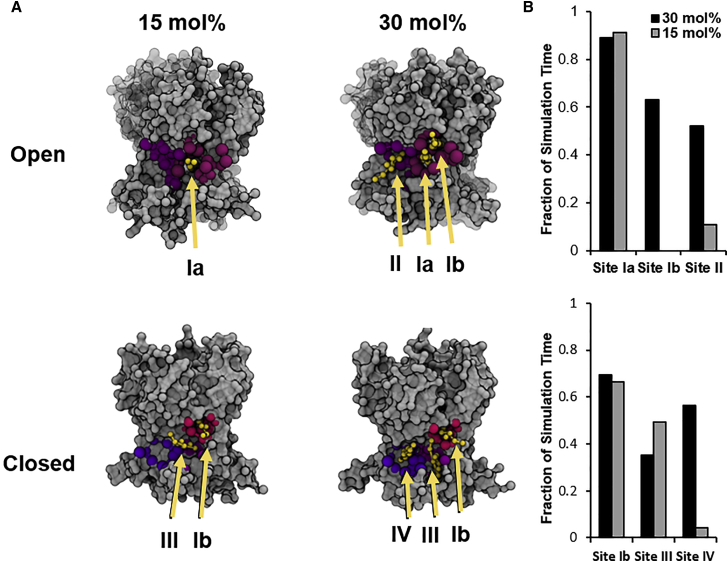
Cholesterol is a major regulator of multiple types of ion channels, but the specific mechanisms and the dynamics of its interactions with the channels are not well understood. Kir2 channels were shown to be sensitive to cholesterol through direct interactions with "cholesterol-sensitive" regions on the channel protein. In this work, we used Martini coarse-grained simulations to analyze the long (μs) timescale dynamics of cholesterol with Kir2.2 channels embedded into a model membrane containing POPC phospholipid with 30 mol% cholesterol. This approach allows us to simulate the dynamic, unbiased migration of cholesterol molecules from the lipid membrane environment to the protein surface of Kir2.2 and explore the favorability of cholesterol interactions at both surface sites and recessed pockets of the channel. We found that the cholesterol environment surrounding Kir channels forms a complex milieu of different short- and long-term interactions, with multiple cholesterol molecules concurrently interacting with the channel. Furthermore, utilizing principles from network theory, we identified four discrete cholesterol-binding sites within the previously identified cholesterol-sensitive region that exist depending on the conformational state of the channel-open or closed. We also discovered that a twofold decrease in the cholesterol level of the membrane, which we found earlier to increase Kir2 activity, results in a site-specific decrease of cholesterol occupancy at these sites in both the open and closed states: cholesterol molecules at the deepest of these discrete sites shows no change in occupancy at different cholesterol levels, whereas the remaining sites showed a marked decrease in occupancy.
Copyright © 2018 Biophysical Society. All rights reserved.
Figures







References
-
- Rosenhouse-Dantsker A., Mehta D., Levitan I. Regulation of ion channels by membrane lipids. Compr. Physiol. 2012;2:31–68. - PubMed
-
- Addona G.H., Sandermann H., Jr., Miller K.W. Low chemical specificity of the nicotinic acetylcholine receptor sterol activation site. Biochim. Biophys. Acta. 2003;1609:177–182. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
