

Regulation of cell surface protease receptor S100A10 by retinoic acid therapy in acute promyelocytic leukemia (APL)☆
- PMID: 30206209
- PMCID: PMC6134137
- DOI: 10.1038/s41419-018-0954-6
Regulation of cell surface protease receptor S100A10 by retinoic acid therapy in acute promyelocytic leukemia (APL)☆
Abstract
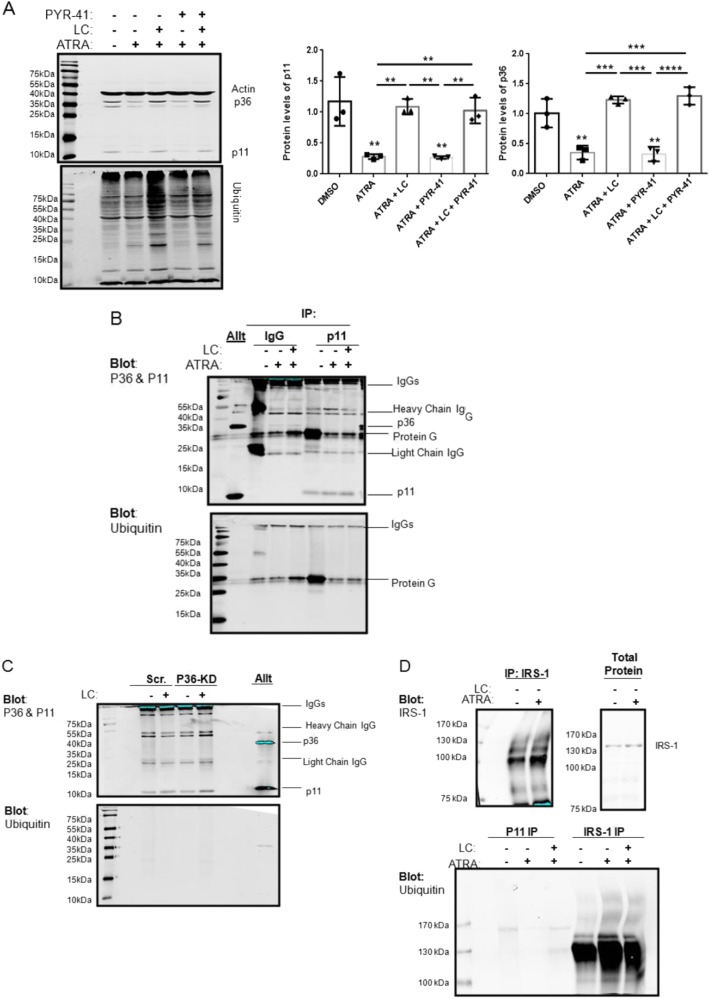
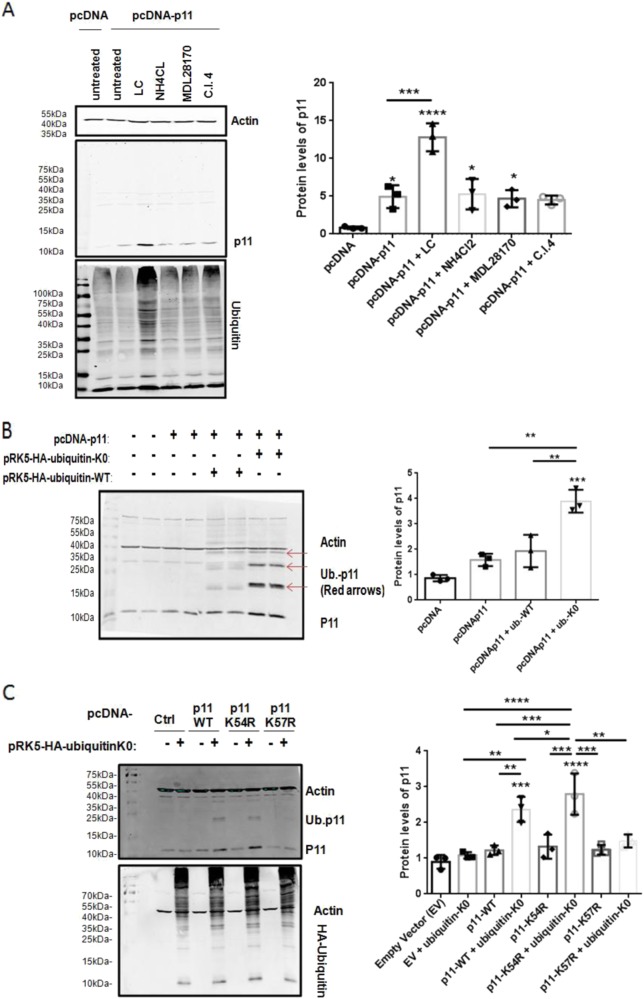
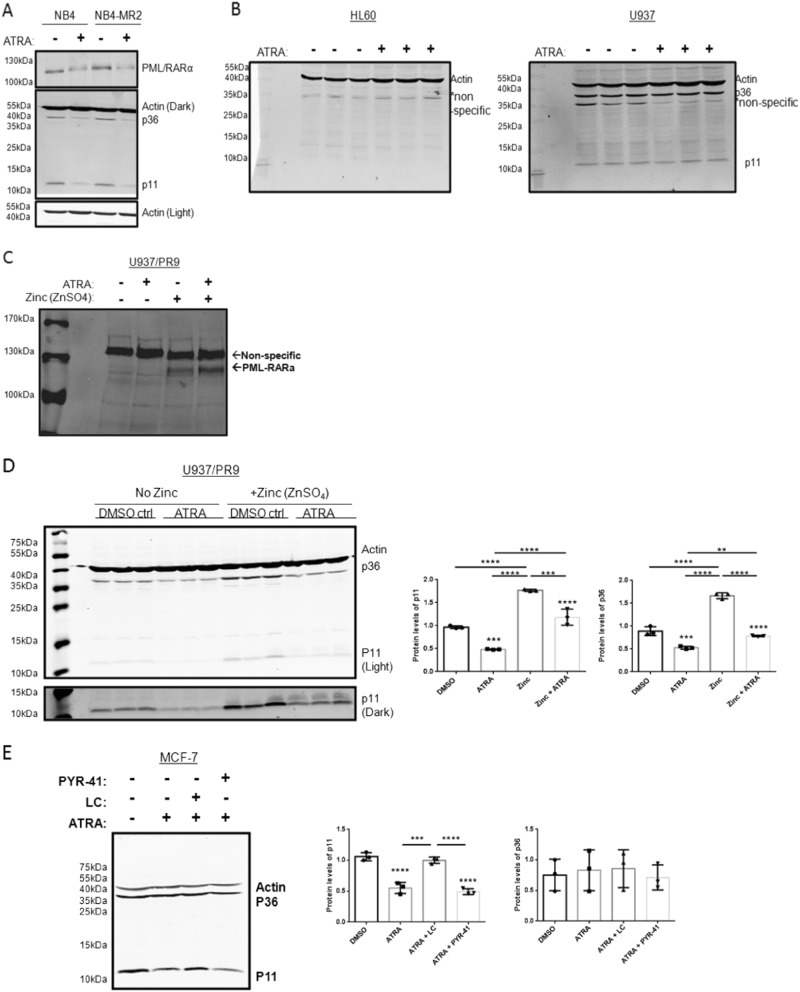
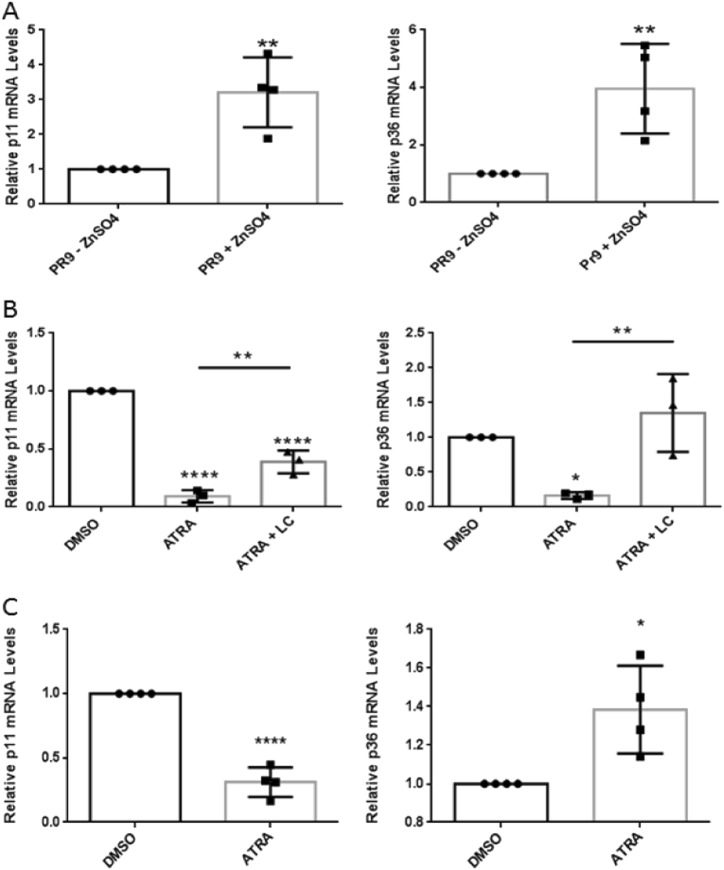
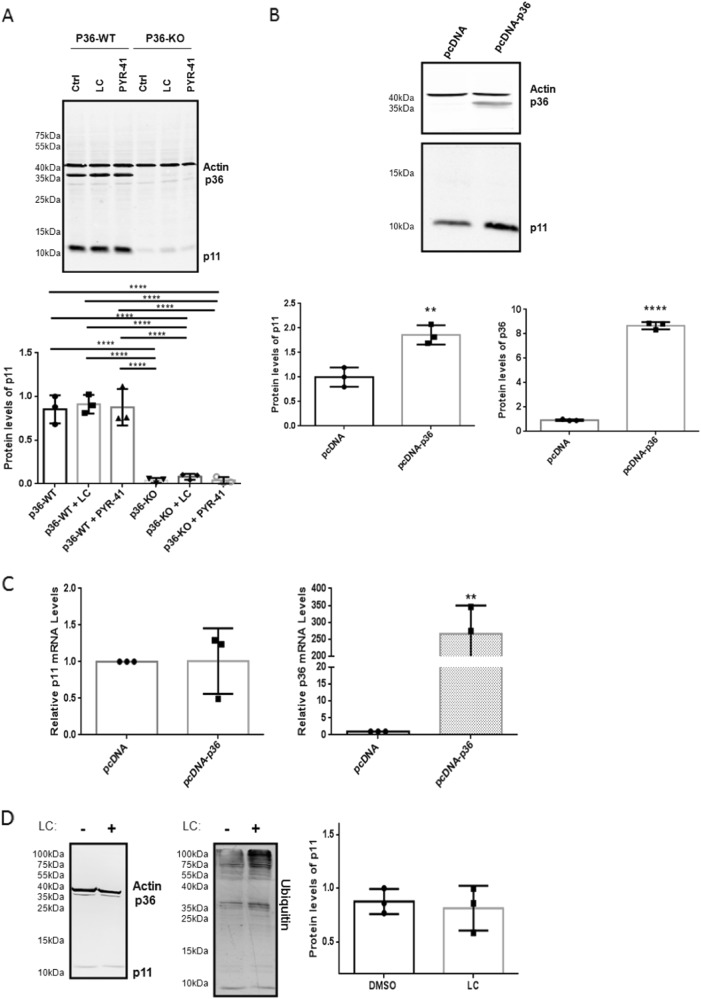
S100A10 (p11), a member of the S100 family of small dimeric EF-hand-type Ca2+-binding proteins, plays a role in a variety of both intracellular and extracellular processes. Previous studies have suggested that p11 is intrinsically unstable and requires binding to annexin A2 (p36) to prevent its rapid ubiquitylation and degradation. Our laboratory has shown that p11 levels are stimulated by the expression of the oncoprotein, PML/RARα. Furthermore, treatment of the APL cell line, NB4 with all-trans retinoic acid (ATRA) causes the rapid loss of p36 and p11 protein. However, the mechanism by which ATRA regulates p11 levels has not been established. Here, we show that the proteasomal inhibitor, lactacystin reversed the ATRA-dependent loss of p11, but did not cause an accumulation of ubiquitylated forms of p11, suggesting that ATRA promotes the proteasomal degradation of p11 in an ubiquitin-independent manner. ATRA treatment of MCF-7 breast cancer cells reduced p11 but not p36 transcript and protein levels, thus indicating that ATRA can regulate p11 levels independently of PML/RARα and p36. Overexpression of p36 upregulated p11 protein but not mRNA levels, indicating that p36 affects p11 post translationally. The forced expression of ubiquitin and p11 in 293 T cells resulted in ubiquitylation of p11 that was blocked by mutagenesis of lysine 57. This study highlights the complex regulation of p11 by retinoid signaling and challenges the hypothesis that ubiquitin-mediated proteasomal degradation of p11 represents a universal mechanism of regulation of this protein.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Annexin A2-S100A10 heterotetramer is upregulated by PML/RARα fusion protein and promotes plasminogen-dependent fibrinolysis and matrix invasion in acute promyelocytic leukemia.Front Med. 2017 Sep;11(3):410-422. doi: 10.1007/s11684-017-0527-6. Epub 2017 Jul 8. Front Med. 2017. PMID: 28687976
-
Regulation of S100A10 by the PML-RAR-α oncoprotein.Blood. 2011 Apr 14;117(15):4095-105. doi: 10.1182/blood-2010-07-298851. Epub 2011 Feb 10. Blood. 2011. PMID: 21310922
-
Endothelial cell annexin A2 regulates polyubiquitination and degradation of its binding partner S100A10/p11.J Biol Chem. 2008 Jul 11;283(28):19192-200. doi: 10.1074/jbc.M800100200. Epub 2008 Apr 23. J Biol Chem. 2008. PMID: 18434302 Free PMC article.
-
Transcription therapy for acute promyelocytic leukaemia.Expert Opin Investig Drugs. 2000 Feb;9(2):329-46. doi: 10.1517/13543784.9.2.329. Expert Opin Investig Drugs. 2000. PMID: 11060680 Review.
-
Novel treatment of acute promyelocytic leukemia: As₂O₃, retinoic acid and retinoid pharmacology.Curr Pharm Biotechnol. 2013;14(9):849-58. doi: 10.2174/1389201015666140113095812. Curr Pharm Biotechnol. 2013. PMID: 24433507 Review.
Cited by
-
Plasmin and Plasminogen System in the Tumor Microenvironment: Implications for Cancer Diagnosis, Prognosis, and Therapy.Cancers (Basel). 2021 Apr 12;13(8):1838. doi: 10.3390/cancers13081838. Cancers (Basel). 2021. PMID: 33921488 Free PMC article. Review.
-
The ANXA2/S100A10 Complex-Regulation of the Oncogenic Plasminogen Receptor.Biomolecules. 2021 Nov 26;11(12):1772. doi: 10.3390/biom11121772. Biomolecules. 2021. PMID: 34944416 Free PMC article. Review.
-
S100A10 Has a Critical Regulatory Function in Mammary Tumor Growth and Metastasis: Insights Using MMTV-PyMT Oncomice and Clinical Patient Sample Analysis.Cancers (Basel). 2020 Dec 7;12(12):3673. doi: 10.3390/cancers12123673. Cancers (Basel). 2020. PMID: 33297495 Free PMC article.
-
Anti-tumour effects of all-trans retinoid acid on serous ovarian cancer.J Exp Clin Cancer Res. 2019 Jan 8;38(1):10. doi: 10.1186/s13046-018-1017-7. J Exp Clin Cancer Res. 2019. PMID: 30621740 Free PMC article.
-
Role of cardiolipins, mitochondria, and autophagy in the differentiation process activated by all-trans retinoic acid in acute promyelocytic leukemia.Cell Death Dis. 2022 Jan 10;13(1):30. doi: 10.1038/s41419-021-04476-z. Cell Death Dis. 2022. PMID: 35013142 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
