

CRISPR/Cas9-Mediated Multiplex Genome Editing of the BnWRKY11 and BnWRKY70 Genes in Brassica napus L
- PMID: 30208656
- PMCID: PMC6163266
- DOI: 10.3390/ijms19092716
CRISPR/Cas9-Mediated Multiplex Genome Editing of the BnWRKY11 and BnWRKY70 Genes in Brassica napus L
Abstract
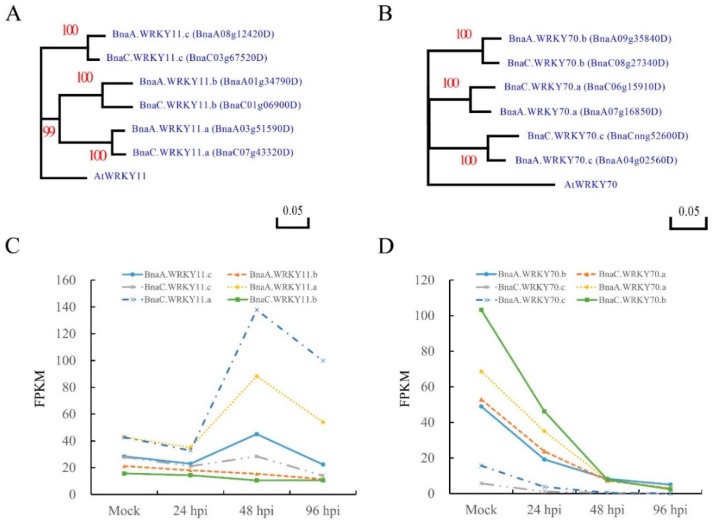
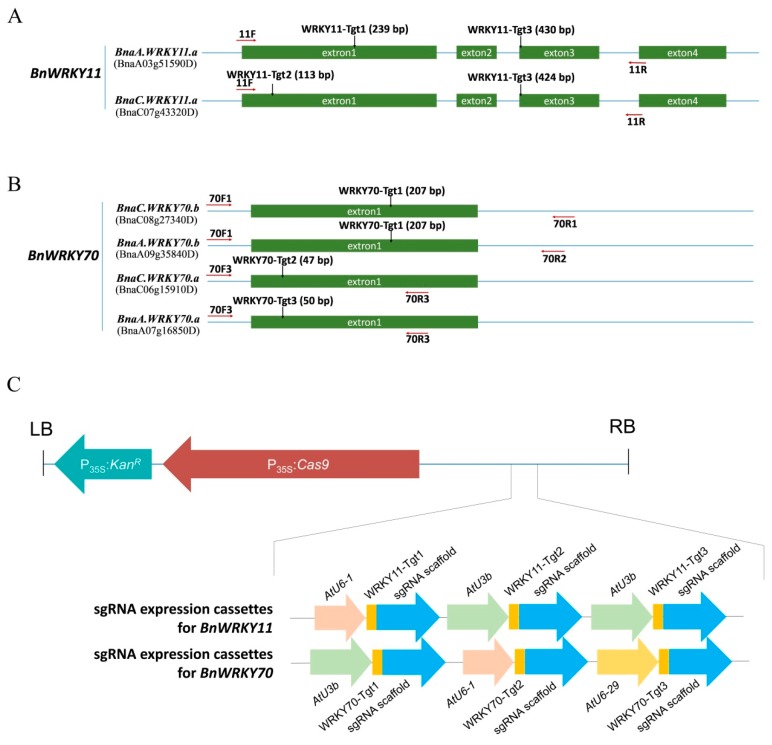
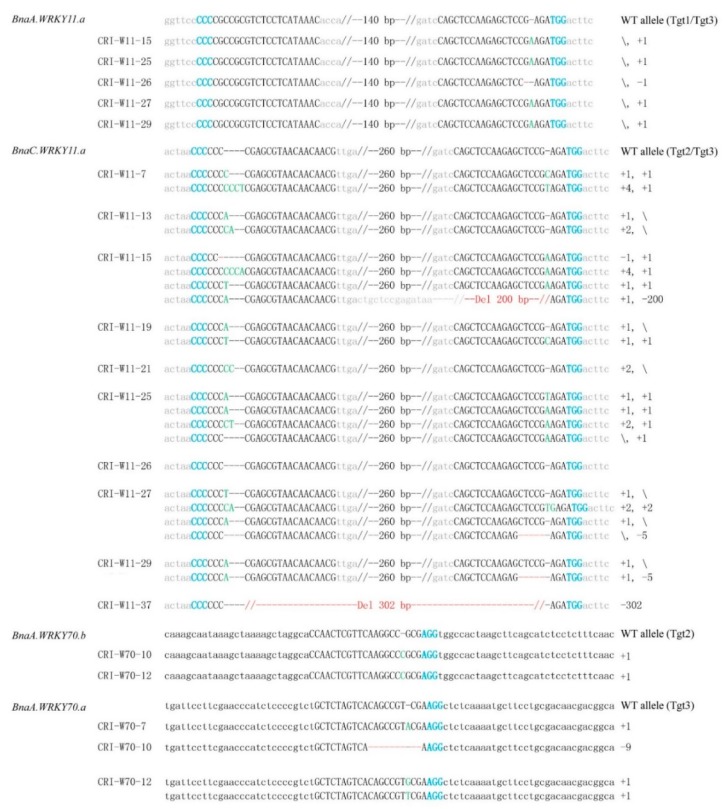
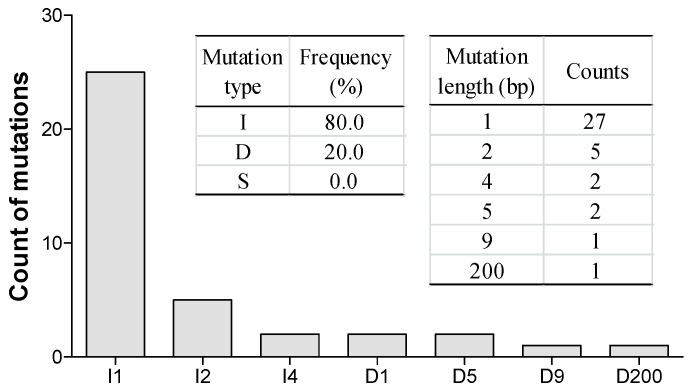
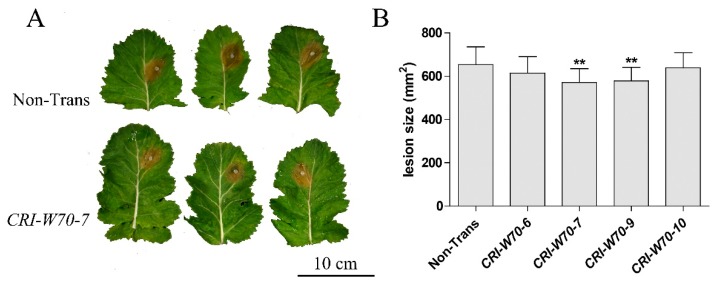
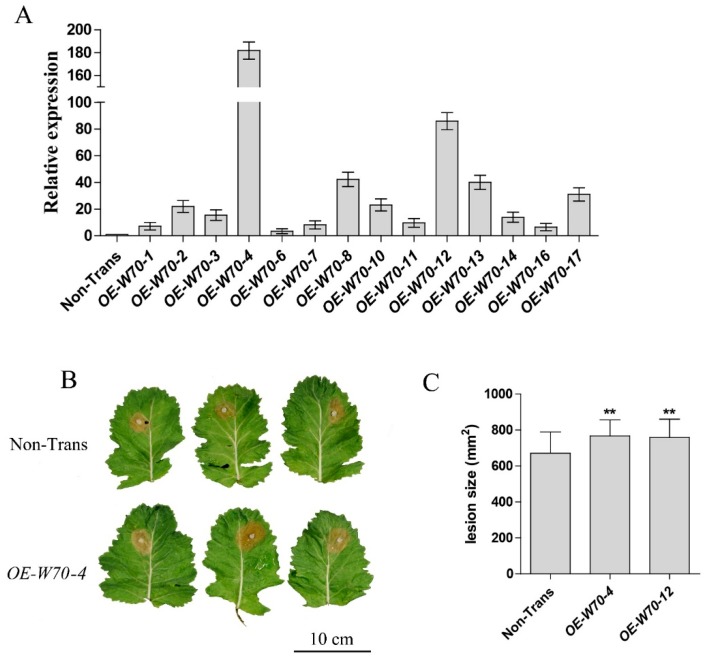
Targeted genome editing is a desirable means of basic science and crop improvement. The clustered, regularly interspaced, palindromic repeat (CRISPR)/Cas9 (CRISPR-associated 9) system is currently the simplest and most commonly used system in targeted genomic editing in plants. Single and multiplex genome editing in plants can be achieved under this system. In Arabidopsis, AtWRKY11 and AtWRKY70 genes were involved in JA- and SA-induced resistance to pathogens, in rapeseed (Brassica napus L.), BnWRKY11 and BnWRKY70 genes were found to be differently expressed after inoculated with the pathogenic fungus, Sclerotinia sclerotiorum (Lib.) de Bary. In this study, two Cas9/sgRNA constructs targeting two copies of BnWRKY11 and four copies of BnWRKY70 were designed to generate BnWRKY11 and BnWRKY70 mutants respectively. As a result, twenty-two BnWRKY11 and eight BnWRKY70 independent transformants (T₀) were obtained, with the mutation ratios of 54.5% (12/22) and 50% (4/8) in BnWRKY11 and BnWRKY70 transformants respectively. Eight and two plants with two copies of mutated BnWRKY11 and BnWRKY70 were obtained respectively. In T₁ generation of each plant examined, new mutations on target genes were detected with high efficiency. The vast majority of BnWRKY70 mutants showed editing in three copies of BnWRKY70 in examined T₁ plants. BnWRKY70 mutants exhibited enhanced resistance to Sclerotinia, while BnWRKY11 mutants showed no significant difference in Sclerotinia resistance when compared to non-transgenic plants. In addition, plants that overexpressed BnWRKY70 showed increased sensitivity when compared to non-transgenic plants. Altogether, our results demonstrated that BnWRKY70 may function as a regulating factor to negatively control the Sclerotinia resistance and CRISPR/Cas9 system could be used to generate germplasm in B. napus with high resistance against Sclerotinia.
Keywords: Brassica napus; CRISPR/Cas9; Mutation pattern; Sclerotinia sclerotiorum; WRKY.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ishino Y., Shinagawa H., Makino K., Amemura M., Nakata A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli and identification of the gene product. J. Bacteriol. 1987;169:5429–5433. doi: 10.1128/jb.169.12.5429-5433.1987. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
