

In vivo CRISPR editing with no detectable genome-wide off-target mutations
- PMID: 30209390
- PMCID: PMC6194229
- DOI: 10.1038/s41586-018-0500-9
In vivo CRISPR editing with no detectable genome-wide off-target mutations
Abstract
CRISPR-Cas genome-editing nucleases hold substantial promise for developing human therapeutic applications1-6 but identifying unwanted off-target mutations is important for clinical translation7. A well-validated method that can reliably identify off-targets in vivo has not been described to date, which means it is currently unclear whether and how frequently these mutations occur. Here we describe 'verification of in vivo off-targets' (VIVO), a highly sensitive strategy that can robustly identify the genome-wide off-target effects of CRISPR-Cas nucleases in vivo. We use VIVO and a guide RNA deliberately designed to be promiscuous to show that CRISPR-Cas nucleases can induce substantial off-target mutations in mouse livers in vivo. More importantly, we also use VIVO to show that appropriately designed guide RNAs can direct efficient in vivo editing in mouse livers with no detectable off-target mutations. VIVO provides a general strategy for defining and quantifying the off-target effects of gene-editing nucleases in whole organisms, thereby providing a blueprint to foster the development of therapeutic strategies that use in vivo gene editing.
Figures
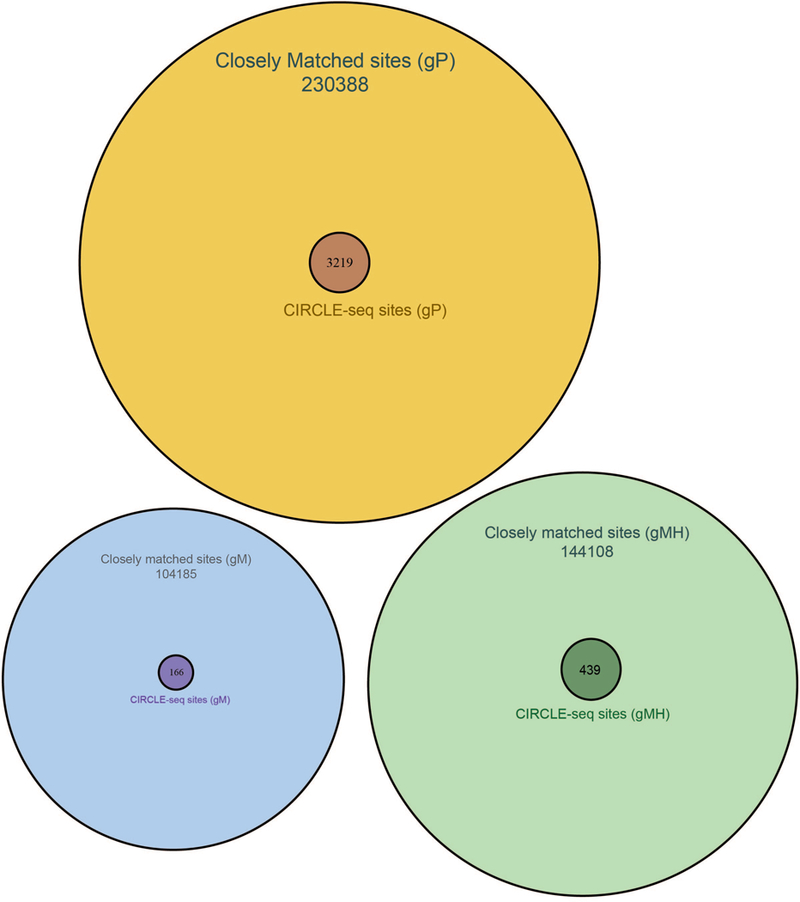
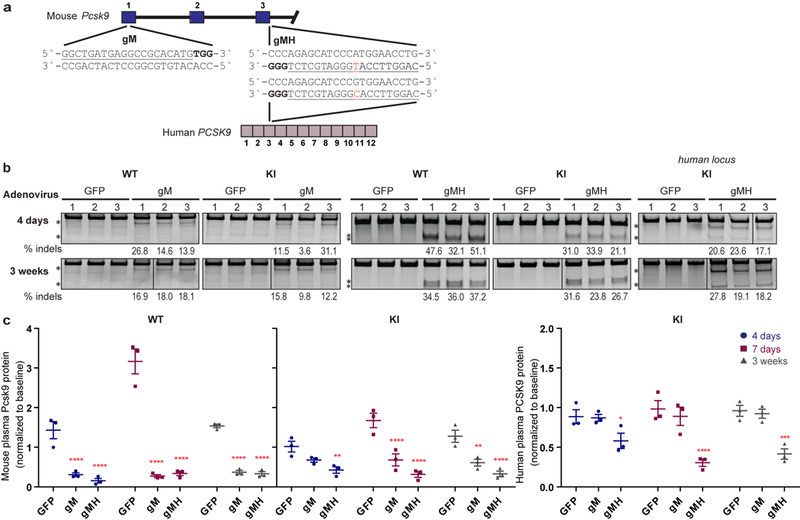
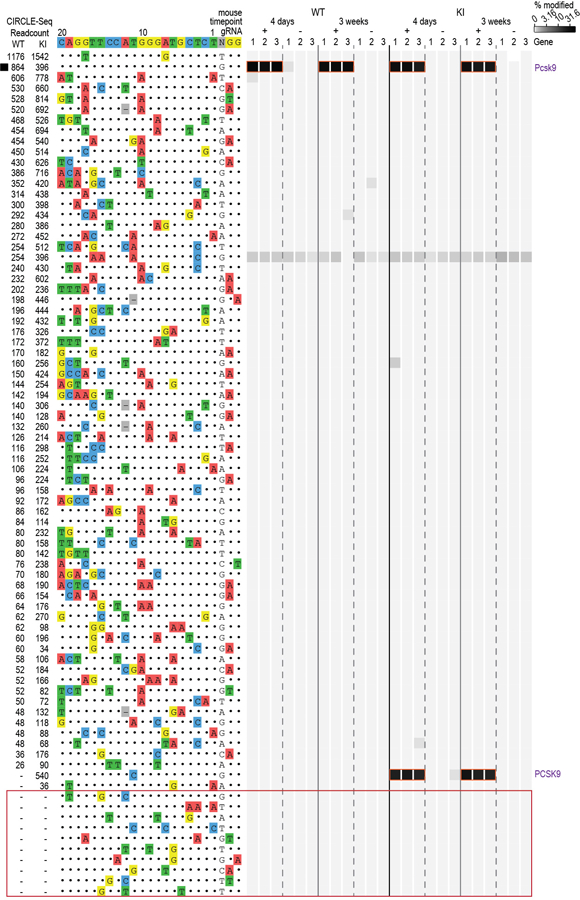
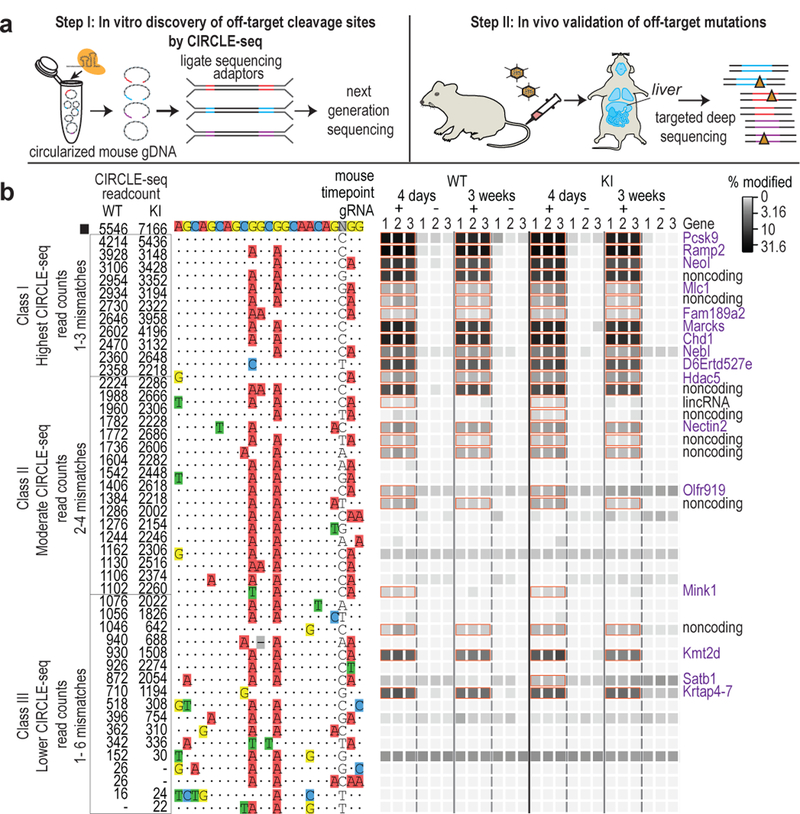

Comment in
-
Challenges and advances of CRISPR-Cas9 genome editing in therapeutics.Cardiovasc Res. 2019 Feb 1;115(2):e12-e14. doi: 10.1093/cvr/cvy300. Cardiovasc Res. 2019. PMID: 30668679 No abstract available.
References
-
- Musunuru K The Hope and Hype of CRISPR-Cas9 Genome Editing: A Review. JAMA cardiology 2, 914–919 (2017). - PubMed
-
- Koo T & Kim JS Therapeutic applications of CRISPR RNA-guided genome editing. Brief Funct Genomics 16, 38–45 (2017). - PubMed
-
- Cornu TI, Mussolino C & Cathomen T Refining strategies to translate genome editing to the clinic. Nat Med 23, 415–423 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
