

Evolution in Neuromodulation-The Differential Roles of Acetylcholine in Higher Order Association vs. Primary Visual Cortices
- PMID: 30210306
- PMCID: PMC6121028
- DOI: 10.3389/fncir.2018.00067
Evolution in Neuromodulation-The Differential Roles of Acetylcholine in Higher Order Association vs. Primary Visual Cortices
Abstract
This review contrasts the neuromodulatory influences of acetylcholine (ACh) on the relatively conserved primary visual cortex (V1), compared to the newly evolved dorsolateral prefrontal association cortex (dlPFC). ACh is critical both for proper circuit development and organization, and for optimal functioning of mature systems in both cortical regions. ACh acts through both nicotinic and muscarinic receptors, which show very different expression profiles in V1 vs. dlPFC, and differing effects on neuronal firing. Cholinergic effects mediate attentional influences in V1, enhancing representation of incoming sensory stimuli. In dlPFC ACh plays a permissive role for network communication. ACh receptor expression and ACh actions in higher visual areas have an intermediate profile between V1 and dlPFC. This changing role of ACh modulation across association cortices may help to illuminate the particular susceptibility of PFC in cognitive disorders, and provide therapeutic targets to strengthen cognition.
Keywords: V1; acetylcholine; cholinergic; muscarinic; neuromodulation; nicotinic; prefrontal cortex.
Figures
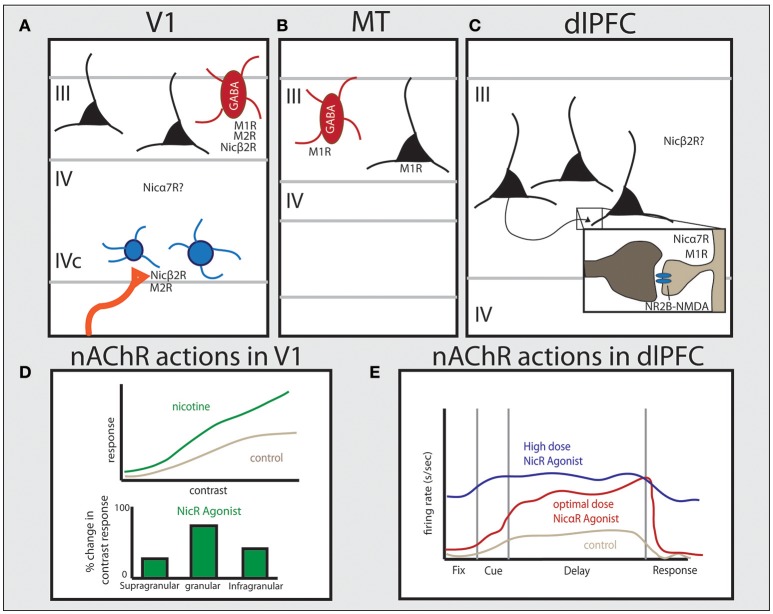
Similar articles
-
Nicotinic α7 receptors enhance NMDA cognitive circuits in dorsolateral prefrontal cortex.Proc Natl Acad Sci U S A. 2013 Jul 16;110(29):12078-83. doi: 10.1073/pnas.1307849110. Epub 2013 Jul 1. Proc Natl Acad Sci U S A. 2013. PMID: 23818597 Free PMC article.
-
Nicotinic α4β2 Cholinergic Receptor Influences on Dorsolateral Prefrontal Cortical Neuronal Firing during a Working Memory Task.J Neurosci. 2017 May 24;37(21):5366-5377. doi: 10.1523/JNEUROSCI.0364-17.2017. Epub 2017 Apr 27. J Neurosci. 2017. PMID: 28450546 Free PMC article.
-
Cholinergic excitation in mouse primary vs. associative cortex: region-specific magnitude and receptor balance.Eur J Neurosci. 2014 Aug;40(4):2608-18. doi: 10.1111/ejn.12622. Epub 2014 May 15. Eur J Neurosci. 2014. PMID: 24827827 Free PMC article.
-
Cholinergic modulation of the medial prefrontal cortex: the role of nicotinic receptors in attention and regulation of neuronal activity.Front Neural Circuits. 2014 Mar 11;8:17. doi: 10.3389/fncir.2014.00017. eCollection 2014. Front Neural Circuits. 2014. PMID: 24653678 Free PMC article. Review.
-
Cholinergic modulation of the cortical neuronal network.Pflugers Arch. 2003 Apr;446(1):17-29. doi: 10.1007/s00424-002-0999-2. Epub 2003 Mar 5. Pflugers Arch. 2003. PMID: 12690458 Review.
Cited by
-
Improving Visual Working Memory with Cholinergic Deep Brain Stimulation.Brain Sci. 2023 Jun 6;13(6):917. doi: 10.3390/brainsci13060917. Brain Sci. 2023. PMID: 37371395 Free PMC article. Review.
-
Morphometric brain organization across the human lifespan reveals increased dispersion linked to cognitive performance.PLoS Biol. 2024 Jun 20;22(6):e3002647. doi: 10.1371/journal.pbio.3002647. eCollection 2024 Jun. PLoS Biol. 2024. PMID: 38900742 Free PMC article.
-
Neurochemistry of Visual Attention.Front Neurosci. 2021 May 5;15:643597. doi: 10.3389/fnins.2021.643597. eCollection 2021. Front Neurosci. 2021. PMID: 34025339 Free PMC article. Review.
-
Synaptogenesis by Cholinergic Stimulation of Astrocytes.Res Sq [Preprint]. 2023 Feb 13:rs.3.rs-2566078. doi: 10.21203/rs.3.rs-2566078/v1. Res Sq. 2023. Update in: Neurochem Res. 2023 Oct;48(10):3212-3227. doi: 10.1007/s11064-023-03979-9. PMID: 36824819 Free PMC article. Updated. Preprint.
-
Associations between genetic variations and global motion perception.Exp Brain Res. 2019 Oct;237(10):2729-2734. doi: 10.1007/s00221-019-05627-7. Epub 2019 Aug 20. Exp Brain Res. 2019. PMID: 31432227
References
-
- Amatrudo J. M., Weaver C. M., Crimins J. L., Hof P. R., Rosene D. L., Luebke J. I. (2012). Influence of highly distinctive structural properties on the excitability of pyramidal neurons in monkey visual and prefrontal cortices. J. Neurosci. 32, 13644–13660. 10.1523/JNEUROSCI.2581-12.2012 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous