

Structure and function of archaeal histones
- PMID: 30212449
- PMCID: PMC6136690
- DOI: 10.1371/journal.pgen.1007582
Structure and function of archaeal histones
Abstract
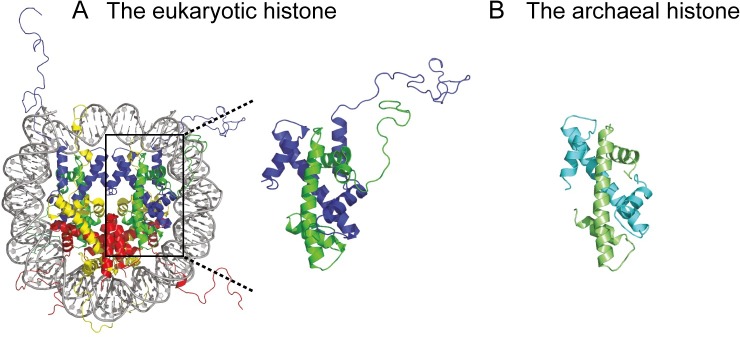
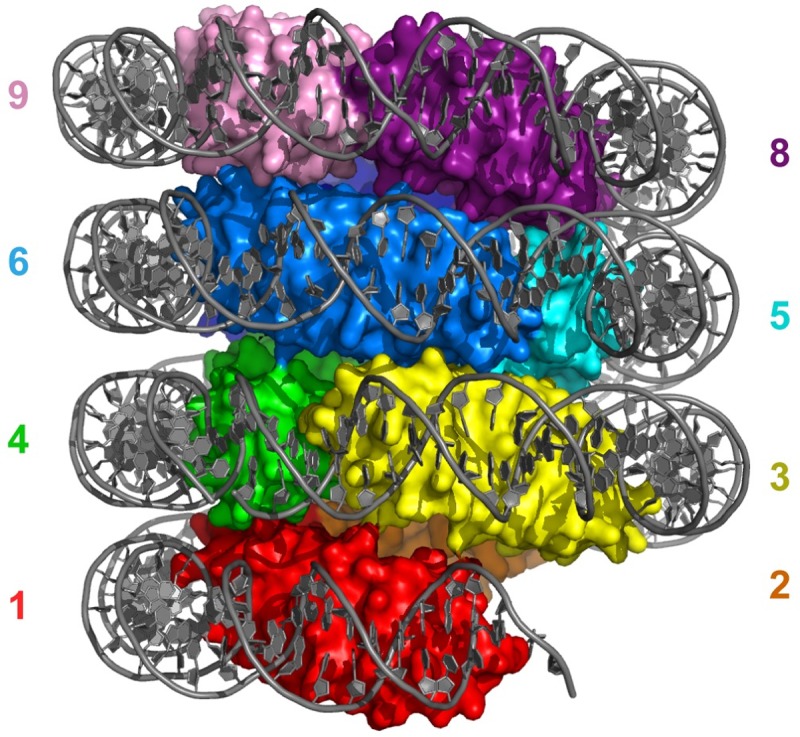
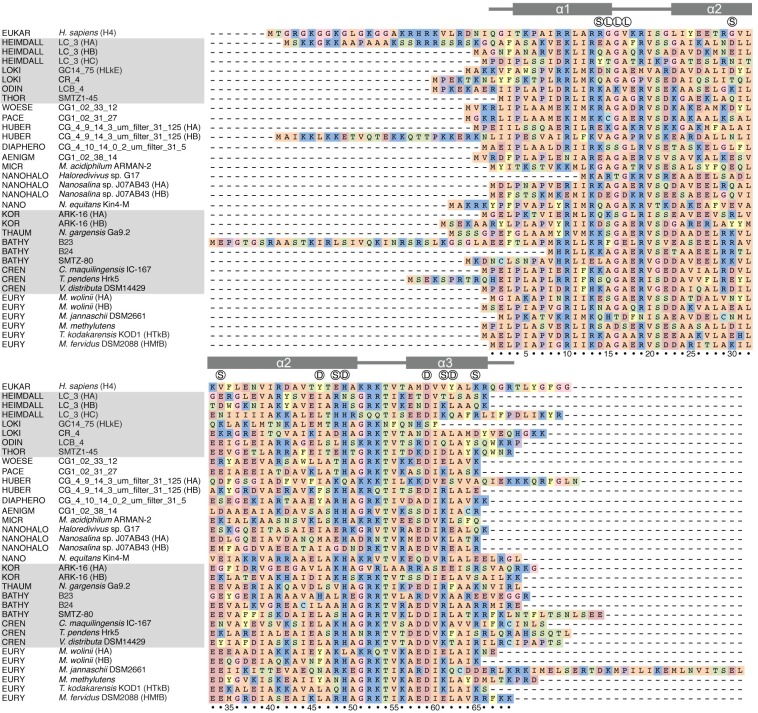
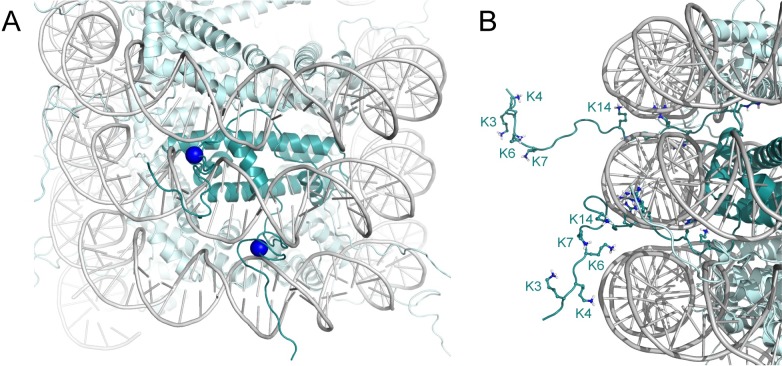
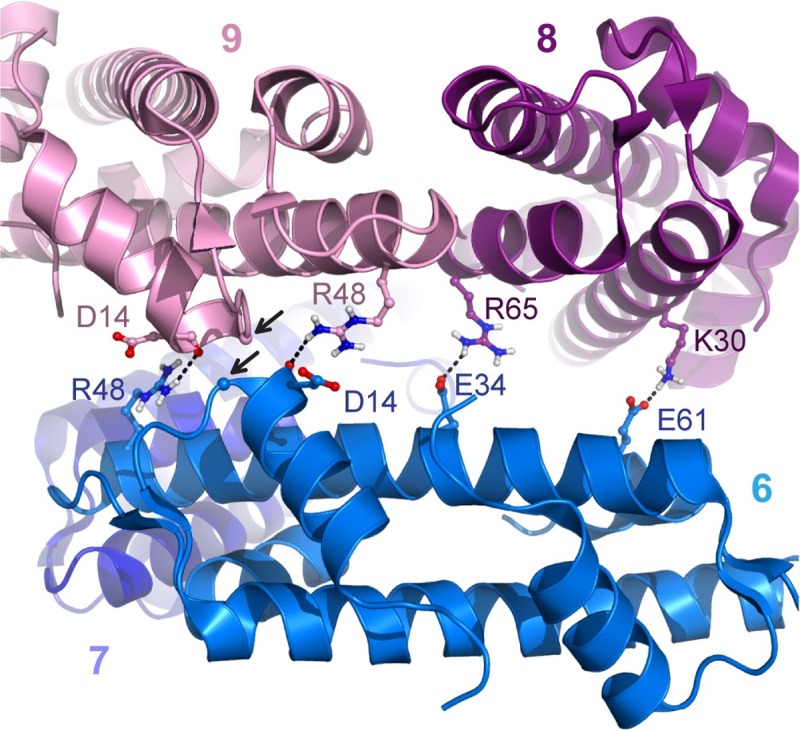
The genomes of all organisms throughout the tree of life are compacted and organized in chromatin by association of chromatin proteins. Eukaryotic genomes encode histones, which are assembled on the genome into octamers, yielding nucleosomes. Post-translational modifications of the histones, which occur mostly on their N-terminal tails, define the functional state of chromatin. Like eukaryotes, most archaeal genomes encode histones, which are believed to be involved in the compaction and organization of their genomes. Instead of discrete multimers, in vivo data suggest assembly of "nucleosomes" of variable size, consisting of multiples of dimers, which are able to induce repression of transcription. Based on these data and a model derived from X-ray crystallography, it was recently proposed that archaeal histones assemble on DNA into "endless" hypernucleosomes. In this review, we discuss the amino acid determinants of hypernucleosome formation and highlight differences with the canonical eukaryotic octamer. We identify archaeal histones differing from the consensus, which are expected to be unable to assemble into hypernucleosomes. Finally, we identify atypical archaeal histones with short N- or C-terminal extensions and C-terminal tails similar to the tails of eukaryotic histones, which are subject to post-translational modification. Based on the expected characteristics of these archaeal histones, we discuss possibilities of involvement of histones in archaeal transcription regulation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
