

Canonical and Non-Canonical Wnt Signaling in Immune Cells
- PMID: 30213499
- PMCID: PMC7367500
- DOI: 10.1016/j.it.2018.08.006
Canonical and Non-Canonical Wnt Signaling in Immune Cells
Abstract
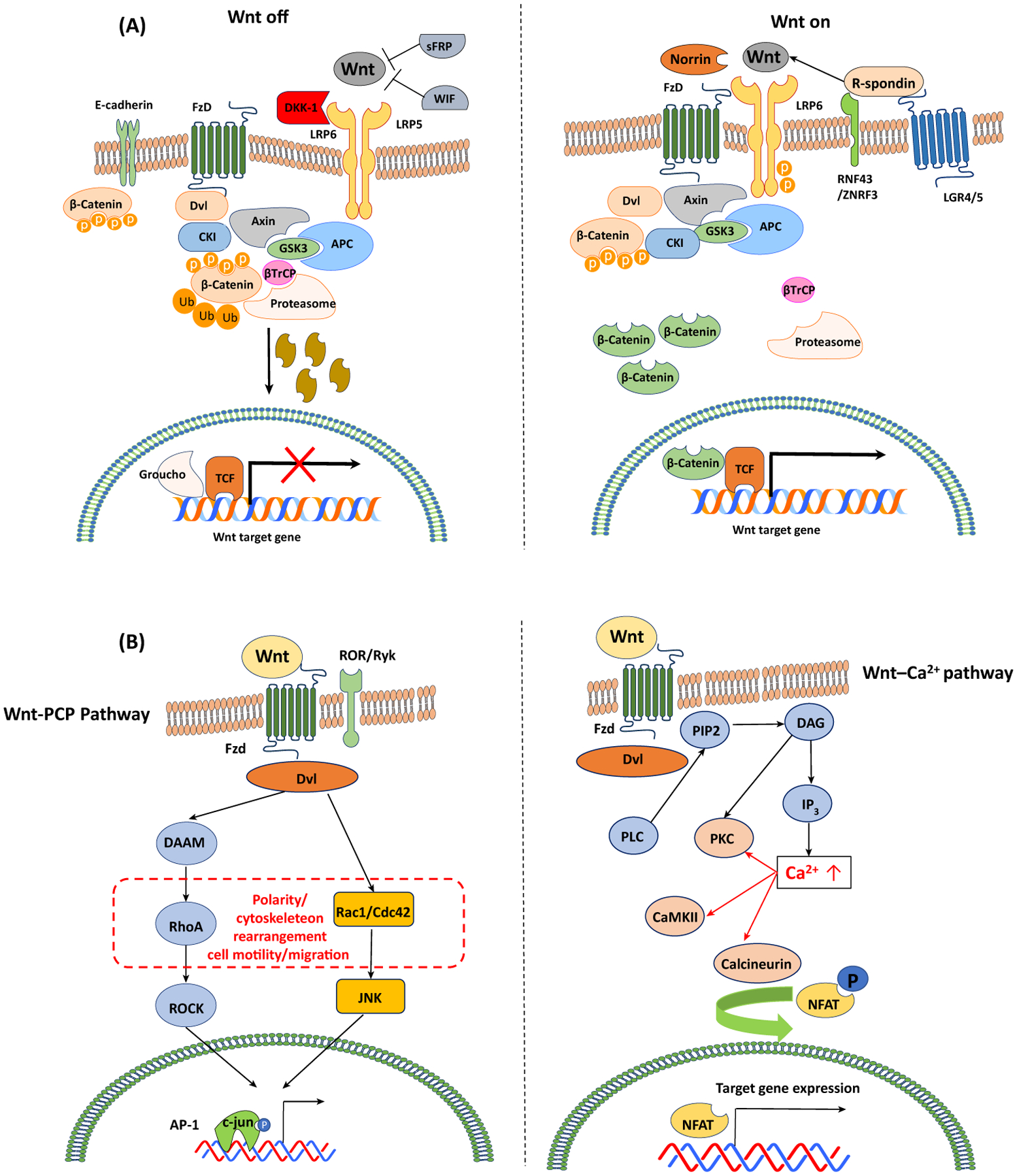
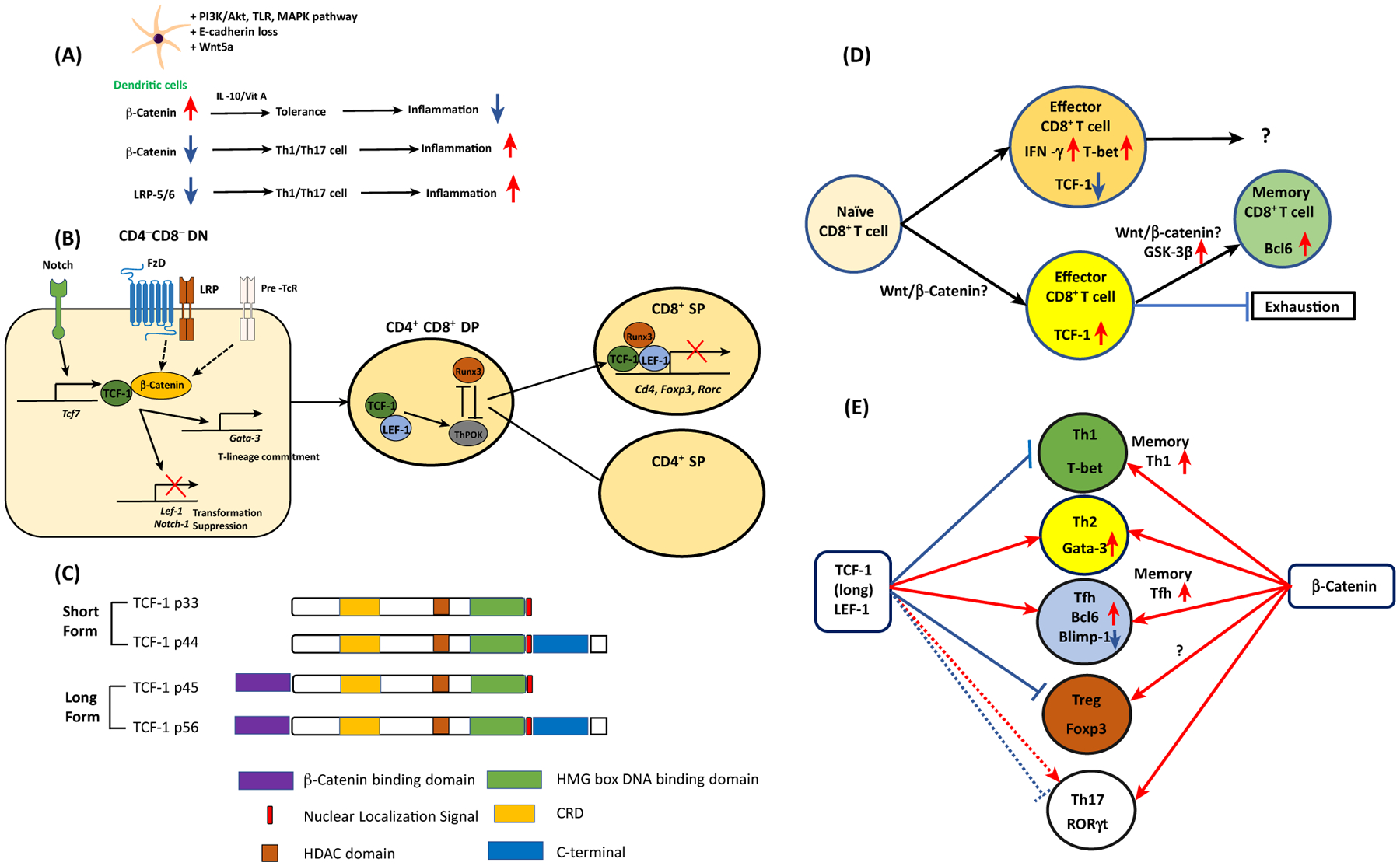
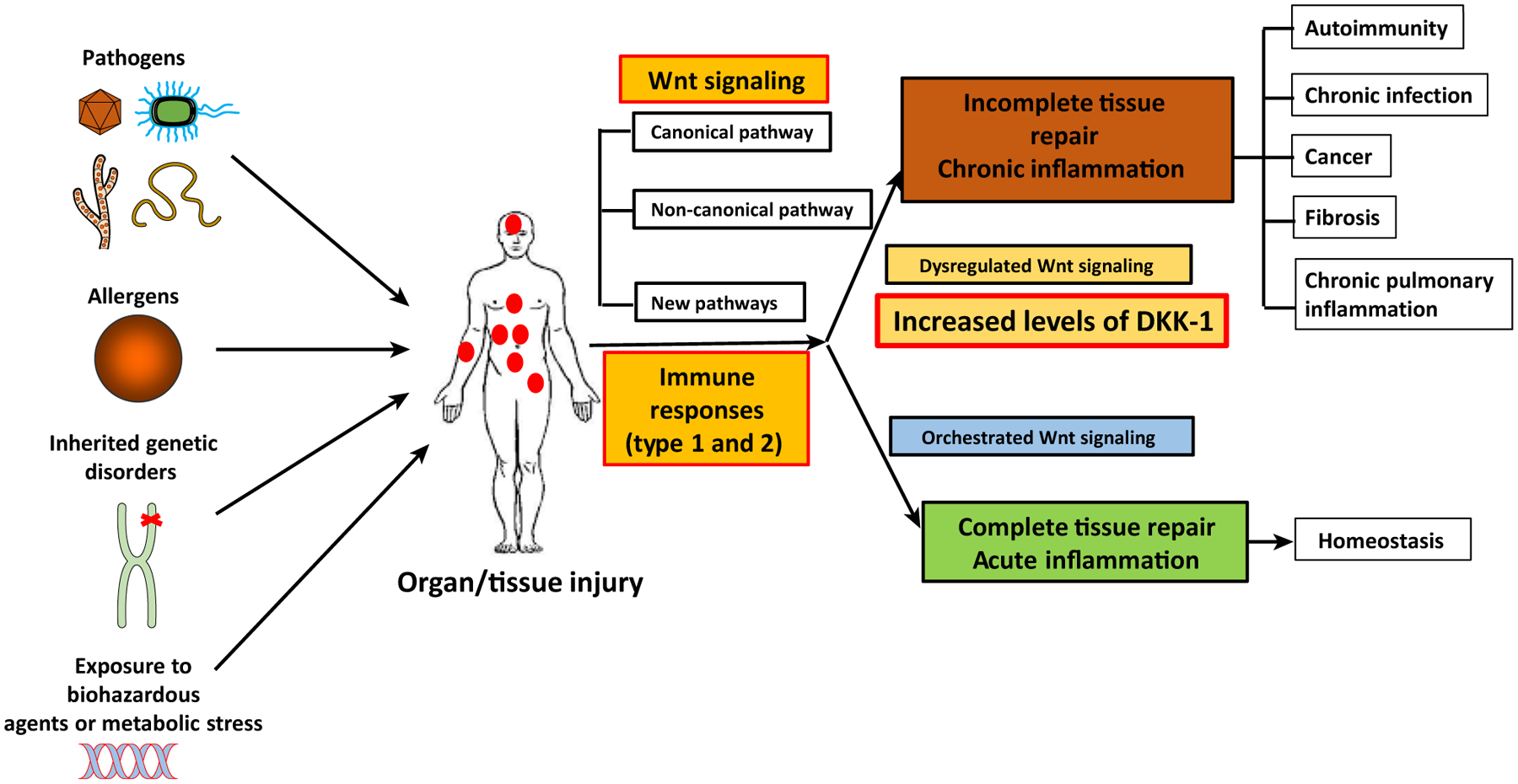
Cell differentiation, proliferation, and death are vital for immune homeostasis. Wnt signaling plays essential roles in processes across species. The roles of Wnt signaling proteins and Wnt ligands have been studied in the past, but the context-dependent mechanisms and functions of these pathways in immune responses remain unclear. Recent findings regarding the role of Wnt ligands and Wnt signaling in immune cells and their immunomodulatory mechanisms suggest that Wnt ligands and signaling are significant in regulating immune responses. We introduce recent key findings and future perspectives on Wnt ligands and their signaling pathways in immune cells as well as the immunological roles and functions of Wnt antagonists.
Copyright © 2018. Published by Elsevier Ltd.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
