

Preserving replication fork integrity and competence via the homologous recombination pathway
- PMID: 30220600
- PMCID: PMC6219450
- DOI: 10.1016/j.dnarep.2018.08.017
Preserving replication fork integrity and competence via the homologous recombination pathway
Abstract
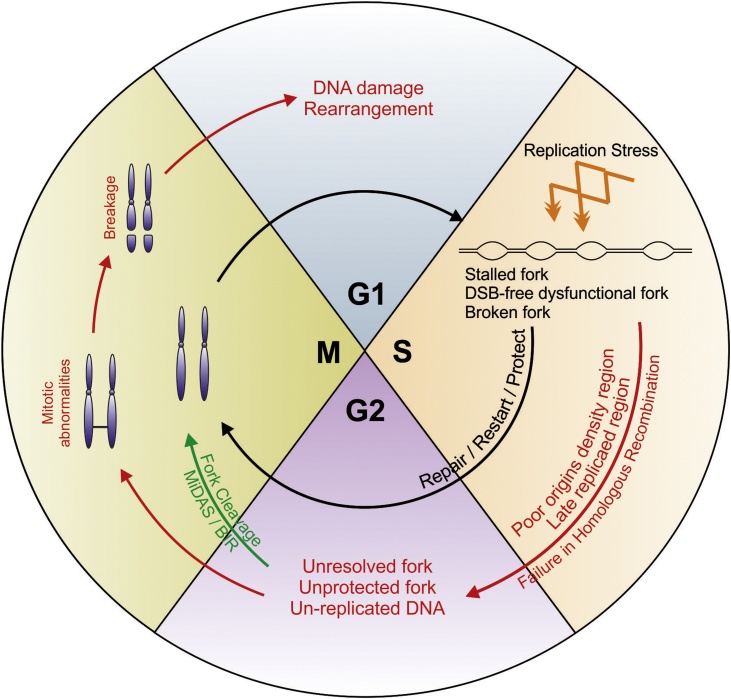
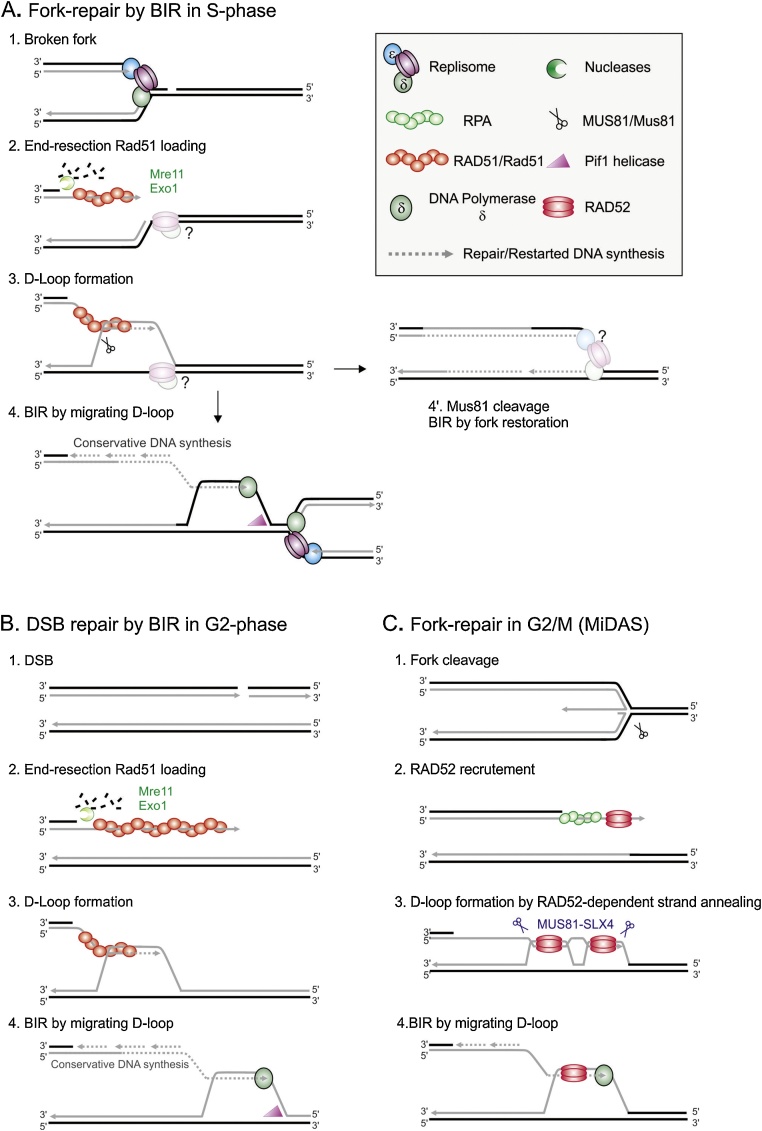
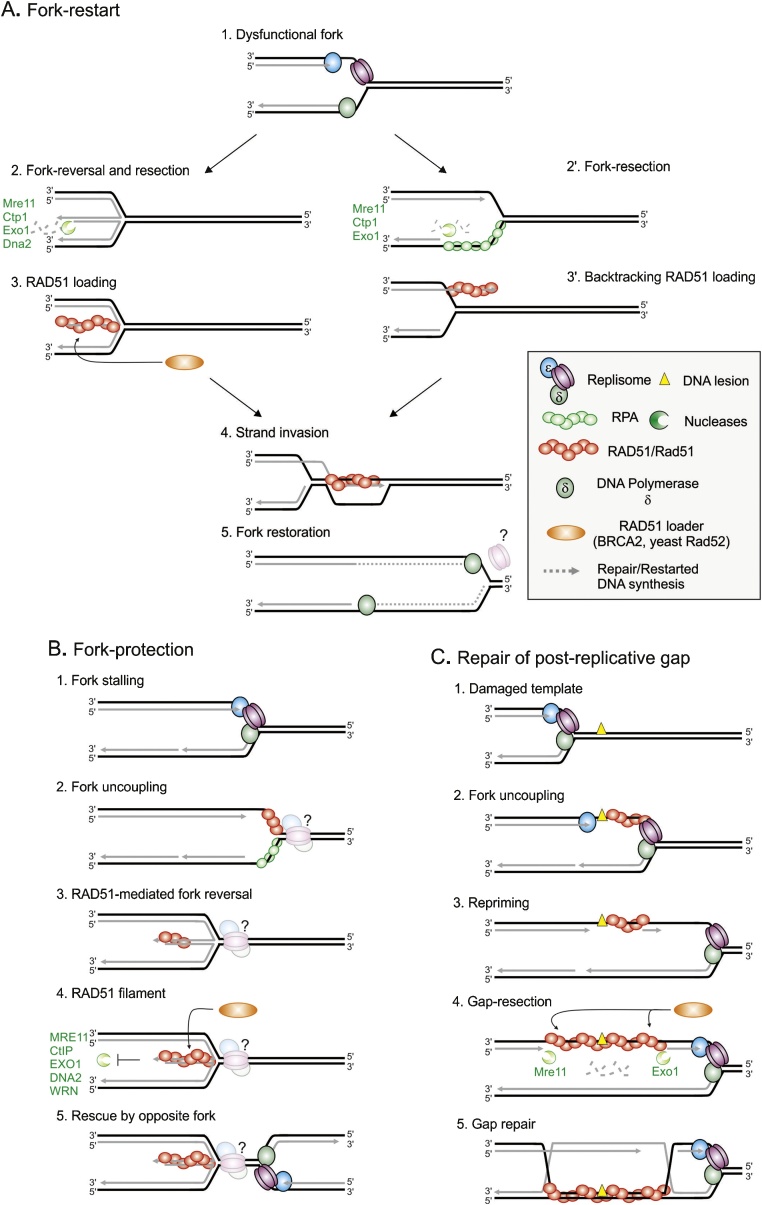
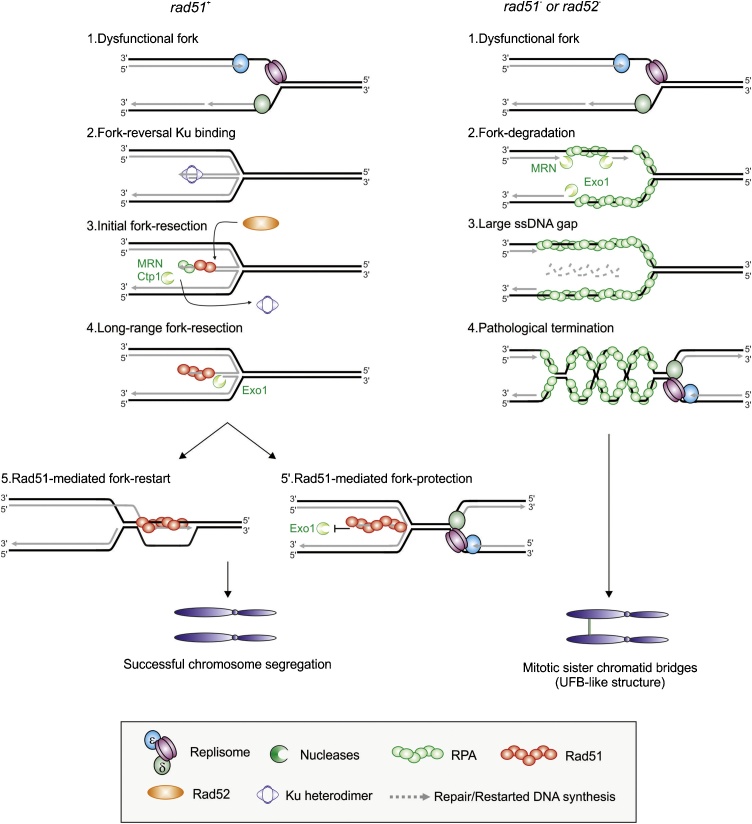
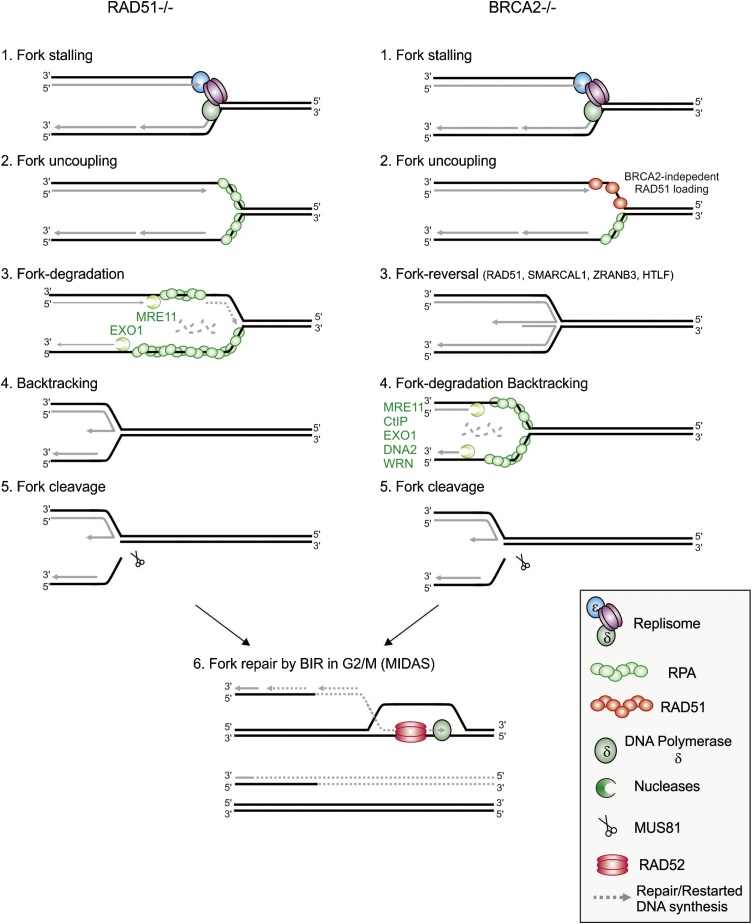
Flaws in the DNA replication process have emerged as a leading driver of genome instability in human diseases. Alteration to replication fork progression is a defining feature of replication stress and the consequent failure to maintain fork integrity and complete genome duplication within a single round of S-phase compromises genetic integrity. This includes increased mutation rates, small and large scale genomic rearrangement and deleterious consequences for the subsequent mitosis that result in the transmission of additional DNA damage to the daughter cells. Therefore, preserving fork integrity and replication competence is an important aspect of how cells respond to replication stress and avoid genetic change. Homologous recombination is a pivotal pathway in the maintenance of genome integrity in the face of replication stress. Here we review our recent understanding of the mechanisms by which homologous recombination acts to protect, restart and repair replication forks. We discuss the dynamics of these genetically distinct functions and their contribution to faithful mitoticsegregation.
Keywords: Fork integrity; Fork restart; Genome instability; Recombination; Replication stress.
Copyright © 2018 The Author(s). Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Técher H., Koundrioukoff S., Nicolas A., Debatisse M. The impact of replication stress on replication dynamics and DNA damage in vertebrate cells. Nat. Rev. Genet. 2017;18:535–550. - PubMed
-
- Gaillard H., García-Muse T., Aguilera A. Replication stress and cancer. Nat. Rev. Cancer. 2015;15:276–289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
