

MiR-24-3p regulates cell proliferation and milk protein synthesis of mammary epithelial cells through menin in dairy cows
- PMID: 30221364
- PMCID: PMC6282567
- DOI: 10.1002/jcp.27017
MiR-24-3p regulates cell proliferation and milk protein synthesis of mammary epithelial cells through menin in dairy cows
Abstract
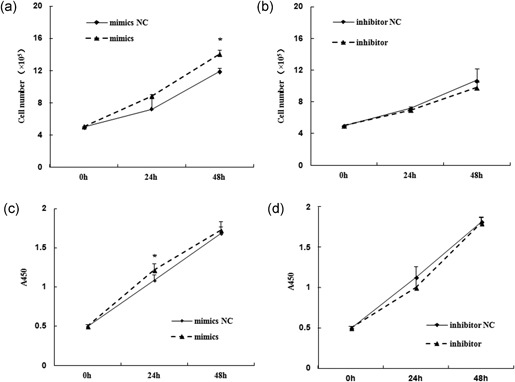
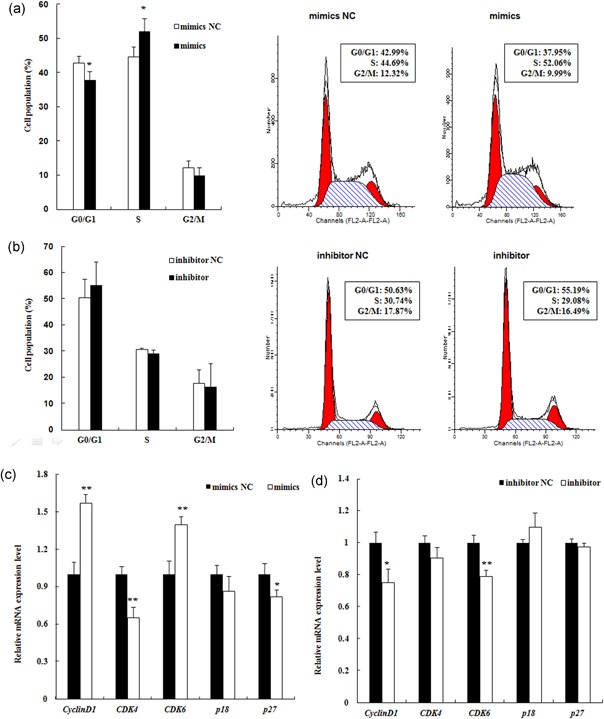
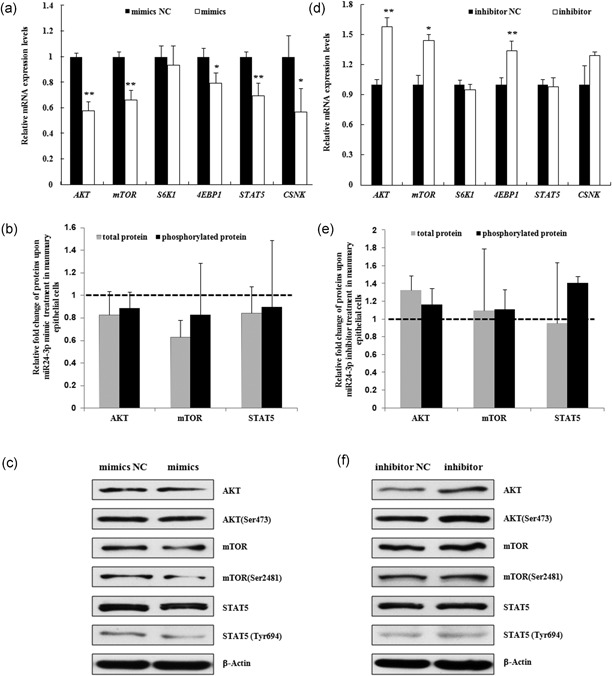
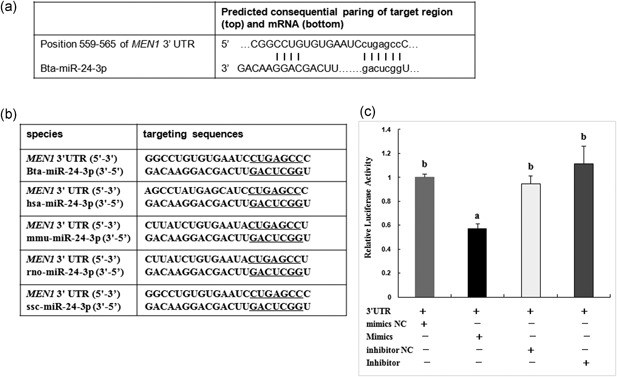
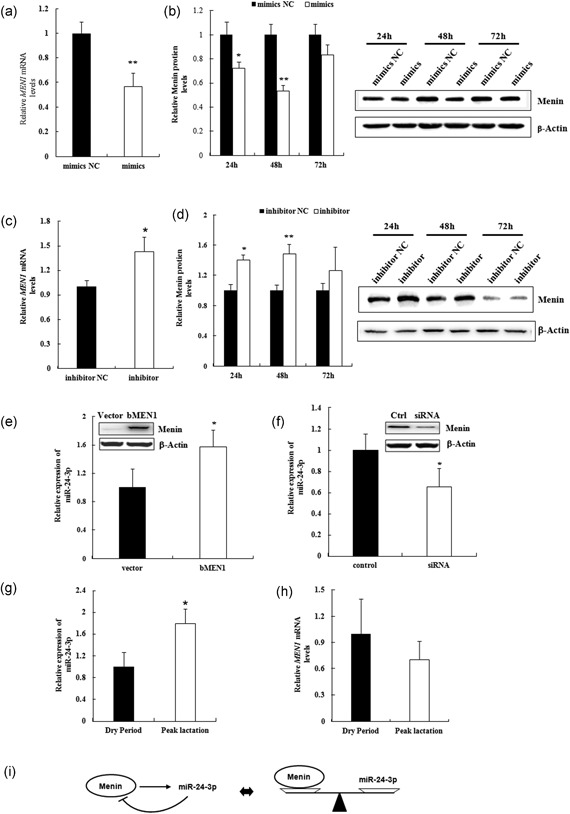
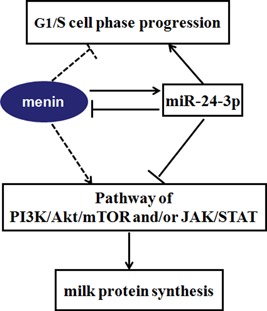
MiR-24-3p, a broadly conserved, small, noncoding RNA, is abundantly expressed in mammary tissue. However, its regulatory role in this tissue remains poorly understood. It was predicted that miR-24-3p targets the 3' untranslated region (3'-UTR) of multiple endocrine neoplasia type 1 (MEN1), an important regulatory factor in mammary tissue. The objective of this study was to investigate the function of miR-24-3p in mammary cells. Using a luciferase assay in mammary epithelial cells (MAC-T), miR-24-3p was confirmed to target the 3'-UTR of MEN1. Furthermore, miR-24-3p negatively regulated the expression of the MEN1 gene and its encoded protein, menin. miR-24-3p enhanced proliferation of MAC-T by promoting G1/S phase progression. MiR-24-3p also regulated the expression of key factors involved in phosphatidylinositol-3-kinase/protein kinase B/mammalian target of rapamycin and Janus kinase/signal transducer and activators of transcription signaling pathways, therefore controlling milk protein synthesis in epithelial cells. Thus, miR-24-3p appears to act on MAC-T by targeting MEN1. The expression of miR-24-3p was controlled by MEN1/menin, indicating a negative feedback loop between miR-24-3p and MEN1/menin. The negatively inhibited expression pattern of miR-24-3p and MEN1 was active in mammary tissues at different lactation stages. The feedback mechanism is a new concept to further understand the lactation cycle of mammary glands and can possibly to be manipulated to improve milk yield and quality.
Keywords: cell proliferation; mammary epithelial cells (MAC-T); miR-24-3p; milk protein synthesis; multiple endocrine neoplasia type 1(MEN1)/menin.
© 2018 The Authors. Journal of Cellular Physiology Published by Wiley Periodicals, Inc.
Figures
References
-
- Appuhamy, J. A. , Nayananjalie, W. A. , England, E. M. , Gerrard, D. E. , Akers, R. M. , & Hanigan, M. D. (2014). Effects of AMP‐activated protein kinase (AMPK) signaling and essential amino acids on mammalian target of rapamycin (mTOR) signaling and protein synthesis rates in mammary cells. Journal of Dairy Science, 97, 419–429. - PubMed
-
- Bartel, D. P. (2004). MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell, 116, 281–297. - PubMed
-
- Bertolino, P. , Tong, W. M. , Galendo, D. , Wang, Z. Q. , & Zhang, C. X. (2003). Heterozygous MEN1 mutant mice develop a range of endocrine tumors mimicking multiple endocrine neoplasia type 1. Molecular Endocrinology, 17(9), 1880–1892. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
