

MAPK and GSK3/ß-TRCP-mediated degradation of the maternal Ets domain transcriptional repressor Yan/Tel controls the spatial expression of nodal in the sea urchin embryo
- PMID: 30222786
- PMCID: PMC6160229
- DOI: 10.1371/journal.pgen.1007621
MAPK and GSK3/ß-TRCP-mediated degradation of the maternal Ets domain transcriptional repressor Yan/Tel controls the spatial expression of nodal in the sea urchin embryo
Abstract
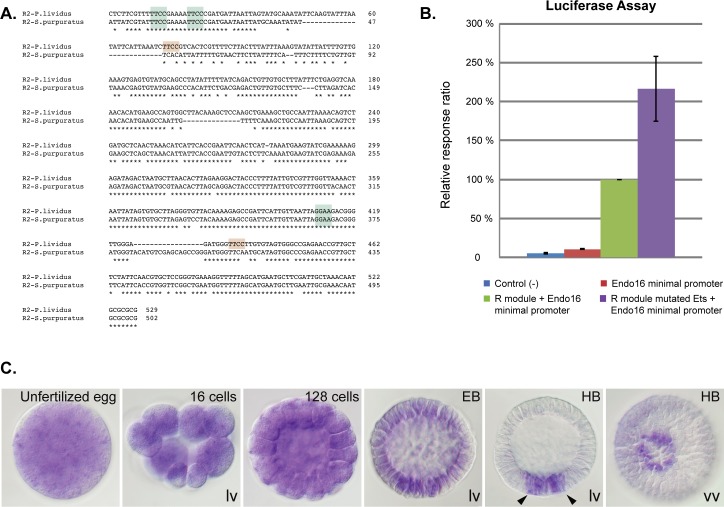
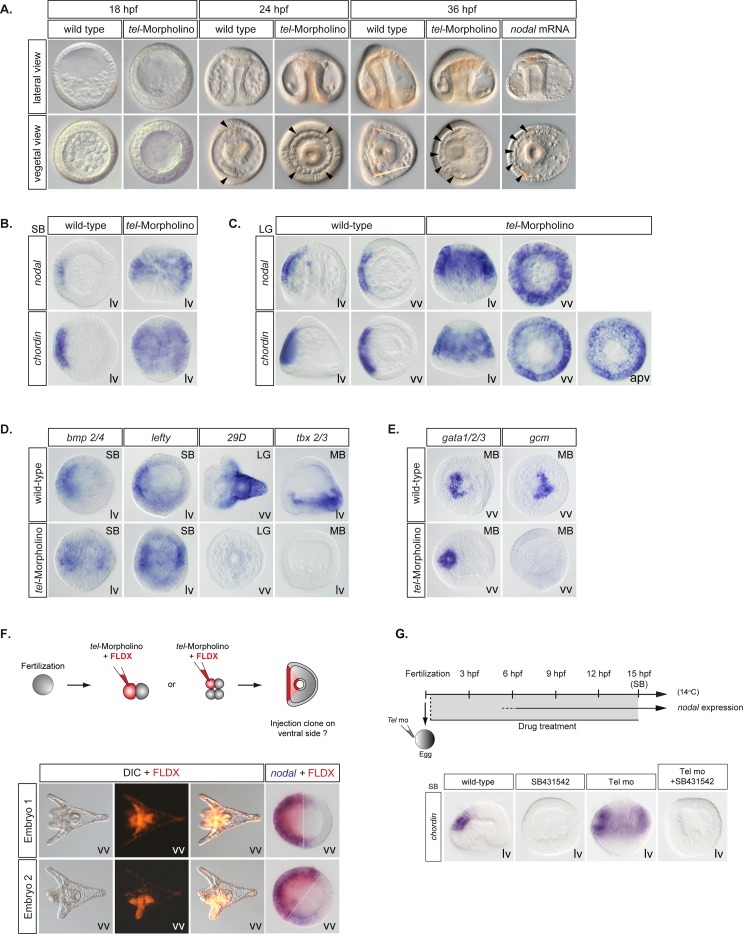
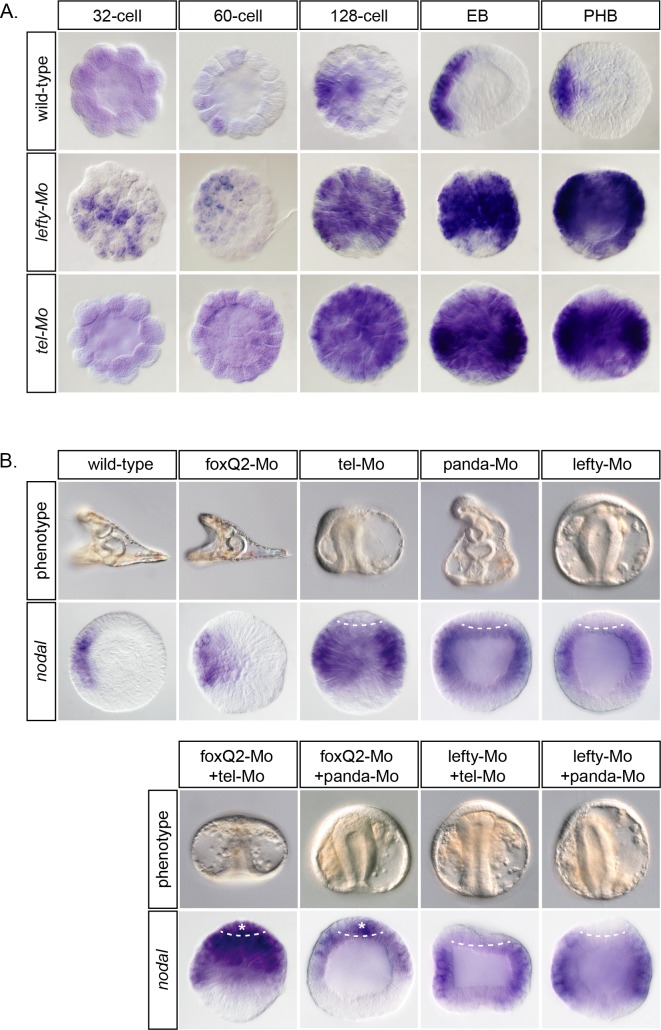
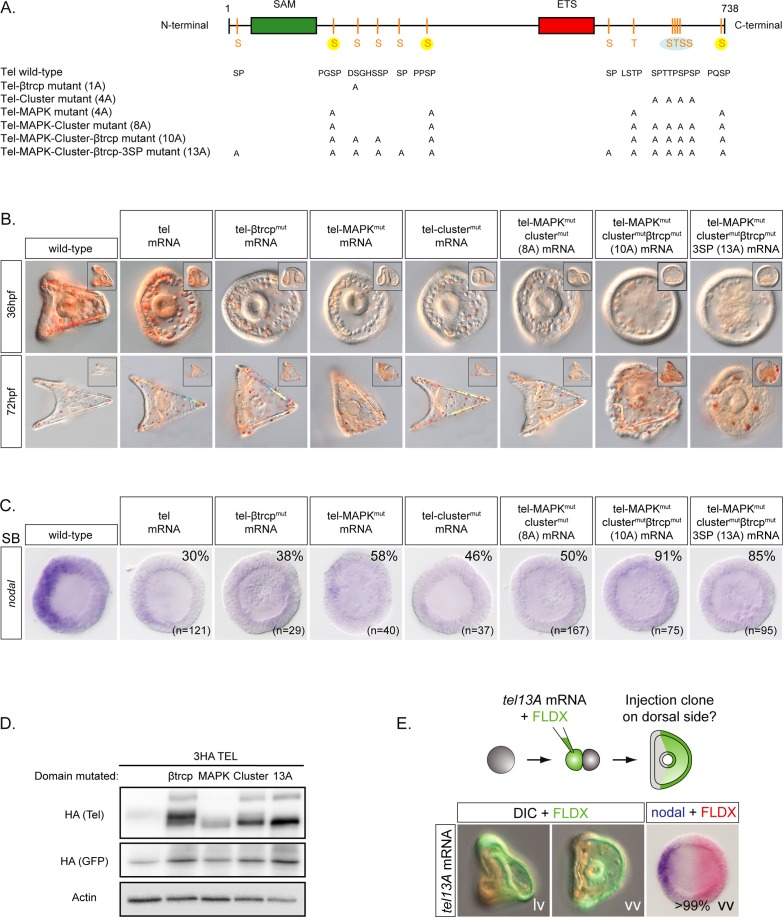
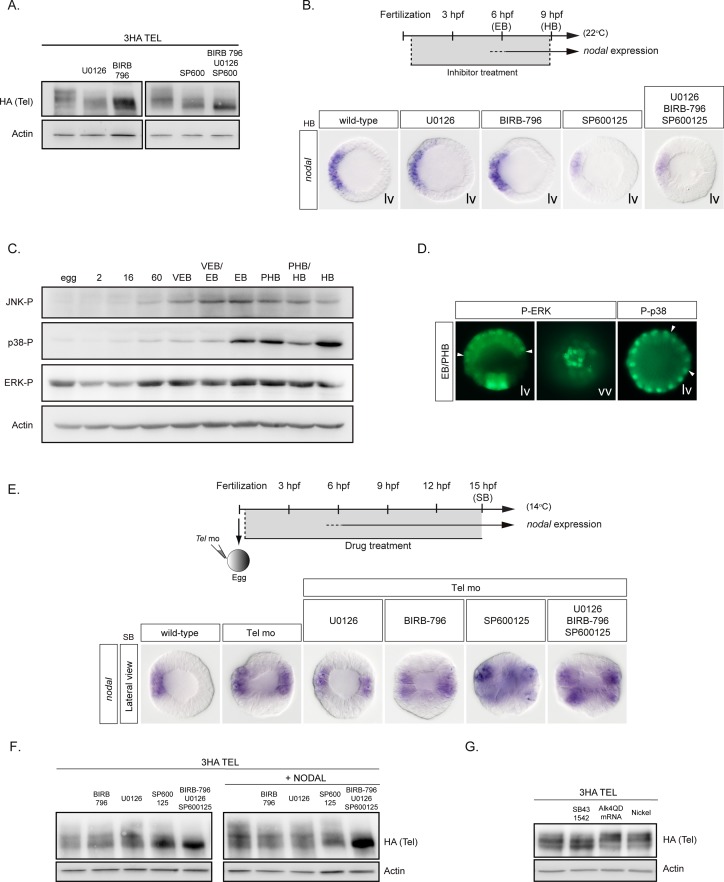
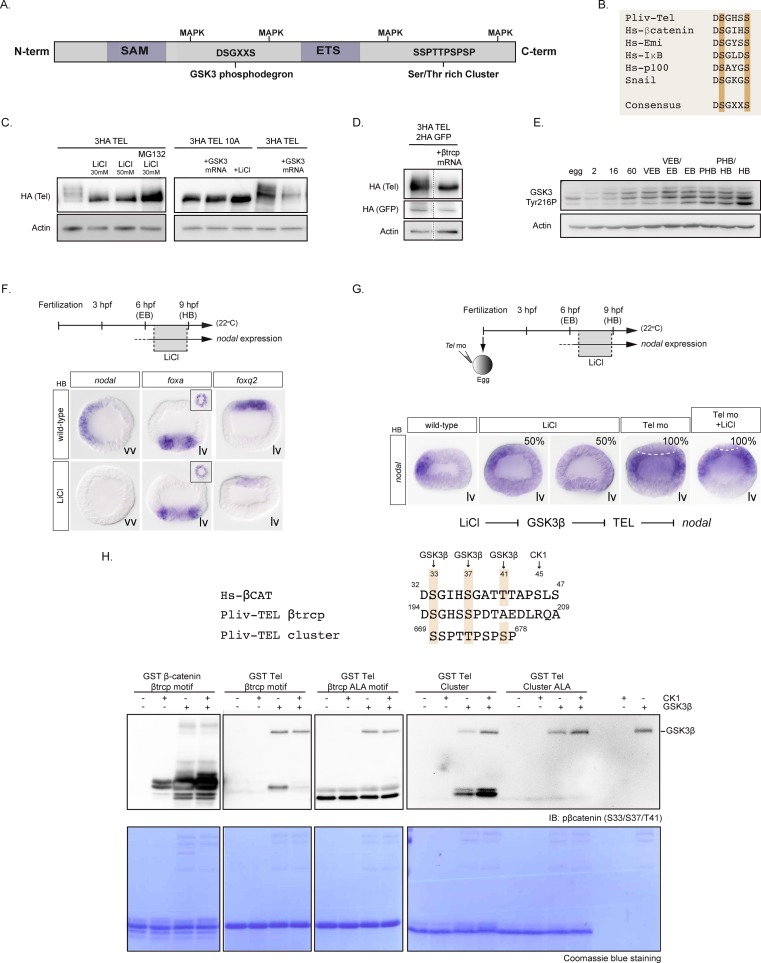
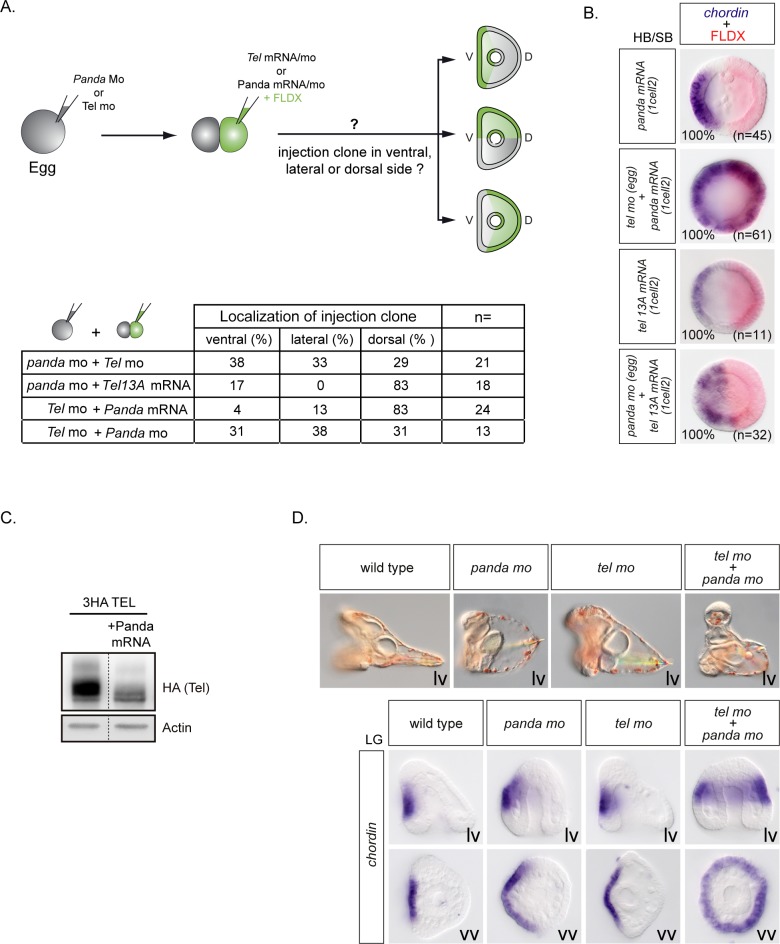
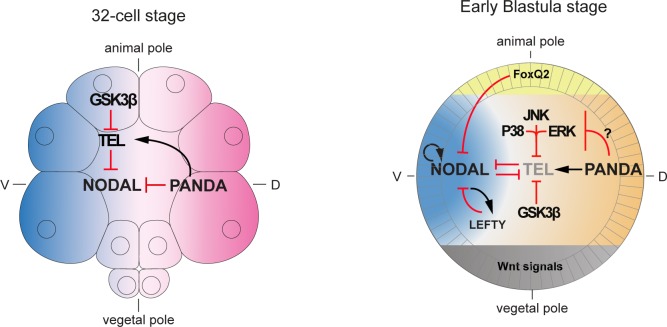
In the sea urchin embryo, specification of the dorsal-ventral axis critically relies on the spatially restricted expression of nodal in the presumptive ventral ectoderm. The ventral restriction of nodal expression requires the activity of the maternal TGF-β ligand Panda but the mechanism by which Panda restricts nodal expression is unknown. Similarly, what initiates expression of nodal in the ectoderm and what are the mechanisms that link patterning along the primary and secondary axes is not well understood. We report that in Paracentrotus lividus, the activity of the maternally expressed ETS-domain transcription factor Yan/Tel is essential for the spatial restriction of nodal. Inhibiting translation of maternal yan/tel mRNA disrupted dorsal-ventral patterning in all germ layers by causing a massive ectopic expression of nodal starting from cleavage stages, mimicking the phenotype caused by inactivation of the maternal Nodal antagonist Panda. We show that like in the fly or in vertebrates, the activity of sea urchin Yan/Tel is regulated by phosphorylation by MAP kinases. However, unlike in the fly or in vertebrates, phosphorylation by GSK3 plays a central role in the regulation Yan/Tel stability in the sea urchin. We show that GSK3 phosphorylates Yan/Tel in vitro at two different sites including a β-TRCP ubiquitin ligase degradation motif and a C-terminal Ser/Thr rich cluster and that phosphorylation of Yan/Tel by GSK3 triggers its degradation by a β-TRCP/proteasome pathway. Finally, we show that, Yan is epistatic to Panda and that the activity of Yan/Tel is required downstream of Panda to restrict nodal expression. Our results identify Yan/Tel as a central regulator of the spatial expression of nodal in Paracentrotus lividus and uncover a key interaction between the gene regulatory networks responsible for patterning the embryo along the dorsal-ventral and animal-vegetal axes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Duboc V, Rottinger E, Besnardeau L, Lepage T. Nodal and BMP2/4 signaling organizes the oral-aboral axis of the sea urchin embryo. Dev Cell. 2004;6(3):397–410. . - PubMed
-
- Meno C, Gritsman K, Ohishi S, Ohfuji Y, Heckscher E, Mochida K, et al. Mouse Lefty2 and zebrafish antivin are feedback inhibitors of nodal signaling during vertebrate gastrulation. Mol Cell. 1999;4(3):287–98. . - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
