

Lignocellulose degradation at the holobiont level: teamwork in a keystone soil invertebrate
- PMID: 30223906
- PMCID: PMC6142342
- DOI: 10.1186/s40168-018-0536-y
Lignocellulose degradation at the holobiont level: teamwork in a keystone soil invertebrate
Abstract
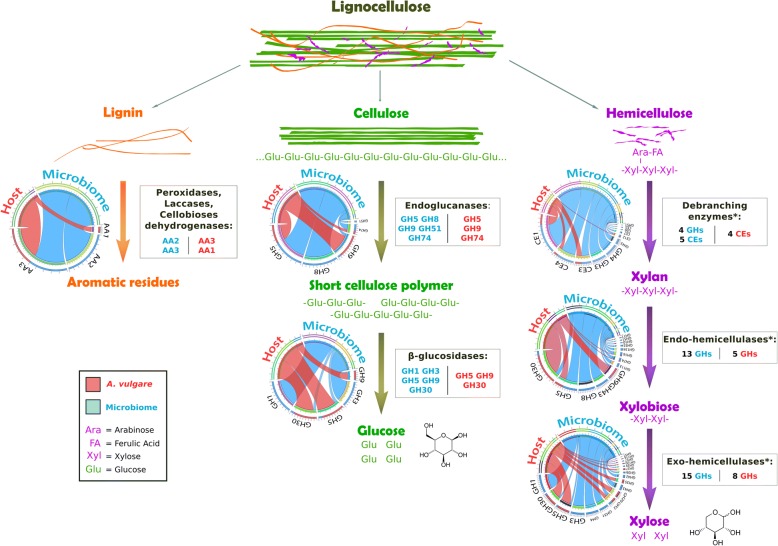
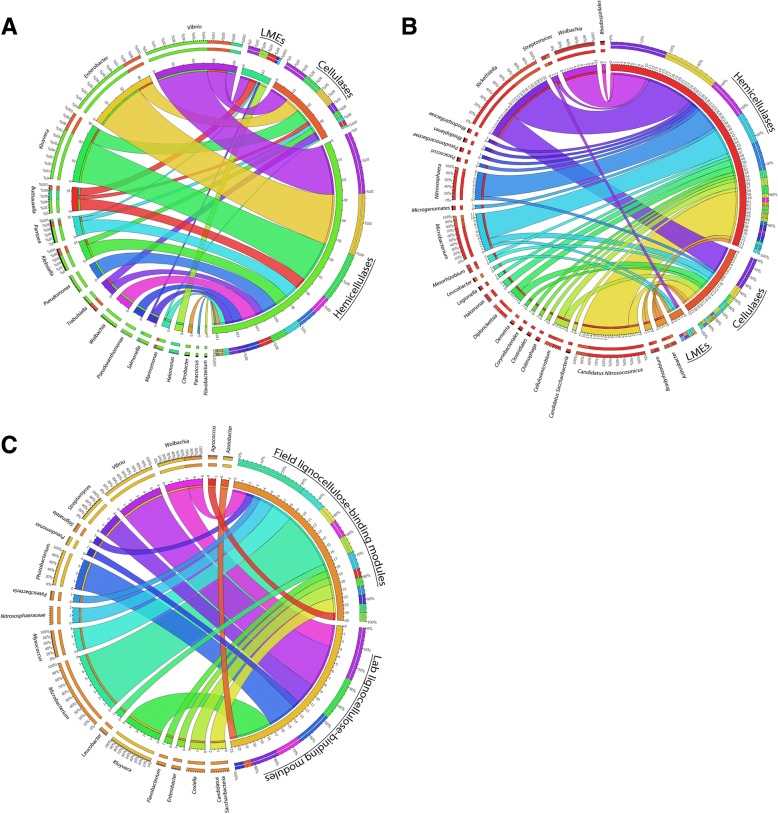
Background: Woodlice are recognized as keystone species in terrestrial ecosystems due to their role in the decomposition of organic matter. Thus, they contribute to lignocellulose degradation and nutrient cycling in the environment together with other macroarthropods. Lignocellulose is the main component of plants and is composed of cellulose, lignin and hemicellulose. Its digestion requires the action of multiple Carbohydrate-Active enZymes (called CAZymes), typically acting together as a cocktail with complementary, synergistic activities and modes of action. Some invertebrates express a few endogenous lignocellulose-degrading enzymes but in most species, an efficient degradation and digestion of lignocellulose can only be achieved through mutualistic associations with endosymbionts. Similar to termites, it has been suspected that several bacterial symbionts may be involved in lignocellulose degradation in terrestrial isopods, by completing the CAZyme repertoire of their hosts.
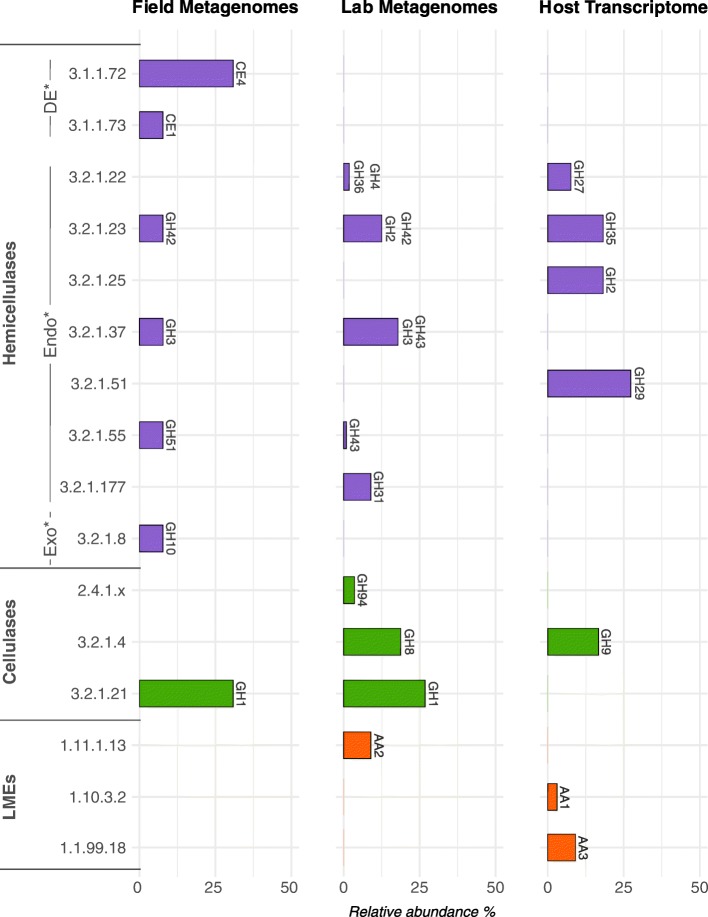
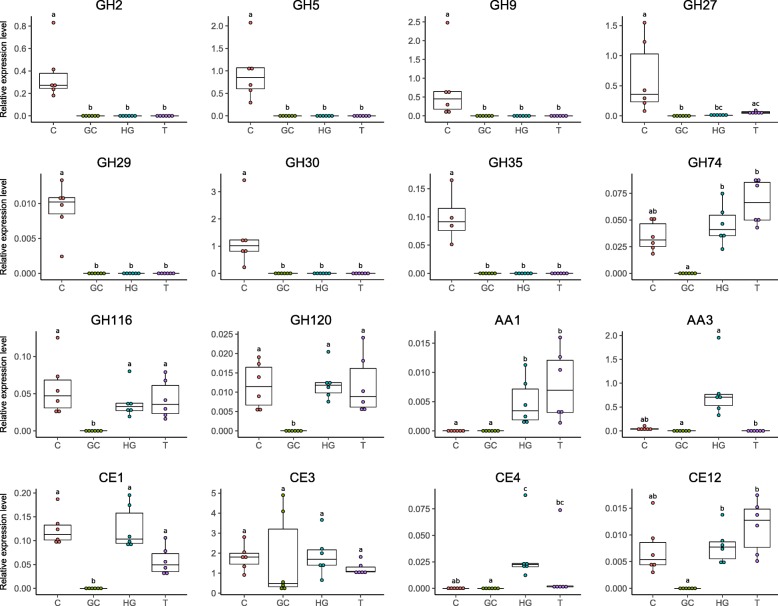
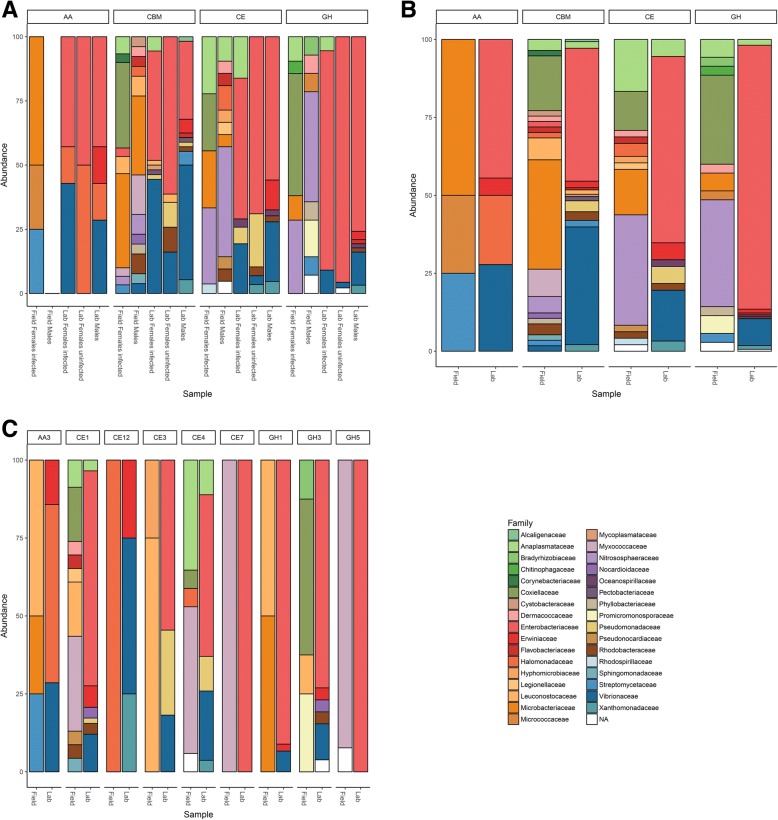
Results: To test this hypothesis, host transcriptomic and microbiome shotgun metagenomic datasets were obtained and investigated from the pill bug Armadillidium vulgare. Many genes of bacterial and archaeal origin coding for CAZymes were identified in the metagenomes of several host tissues and the gut content of specimens from both laboratory lineages and a natural population of A. vulgare. Some of them may be involved in the degradation of cellulose, hemicellulose, and lignin. Reconstructing a lignocellulose-degrading microbial community based on the prokaryotic taxa contributing relevant CAZymes revealed two taxonomically distinct but functionally redundant microbial communities depending on host origin. In parallel, endogenous CAZymes were identified from the transcriptome of the host and their expression in digestive tissues was demonstrated by RT-qPCR, demonstrating a complementary enzyme repertoire for lignocellulose degradation from both the host and the microbiome in A. vulgare.
Conclusions: Our results provide new insights into the role of the microbiome in the evolution of terrestrial isopods and their adaptive radiation in terrestrial habitats.
Keywords: CAZyme; Holobiont; Host–symbiont interactions; Isopods; Microbiome; RT-qPCR; Transcriptome.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Swift M, Heal O, Anderson J. Decomposition in terrestrial ecosystems. Oxford: Blackwell Scientific Publications; 1979.
-
- Hättenschwiler S, Tiunov AV, Scheu S. Biodiversity and litter decomposition in terrestrial ecosystems. Annu Rev Ecol Evol Syst. 2005;36:191–218. doi: 10.1146/annurev.ecolsys.36.112904.151932. - DOI
-
- David JF. The role of litter-feeding macroarthropods in decomposition processes: a reappraisal of common views. Soil Biol Biochem. 2014;76:109–118. doi: 10.1016/j.soilbio.2014.05.009. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
