

Regularizing firing patterns of rat subthalamic neurons ameliorates parkinsonian motor deficits
- PMID: 30226827
- PMCID: PMC6264721
- DOI: 10.1172/JCI99986
Regularizing firing patterns of rat subthalamic neurons ameliorates parkinsonian motor deficits
Abstract
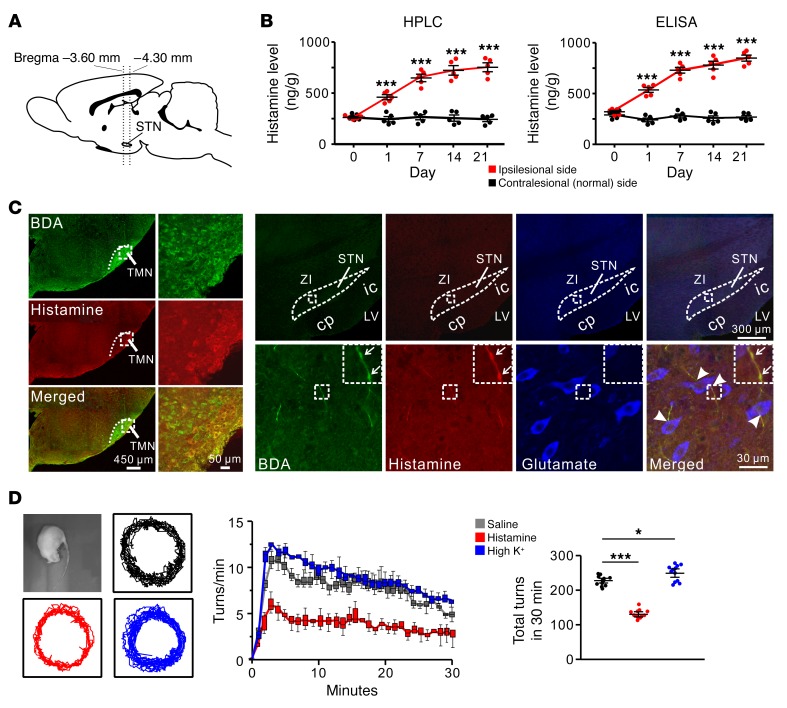
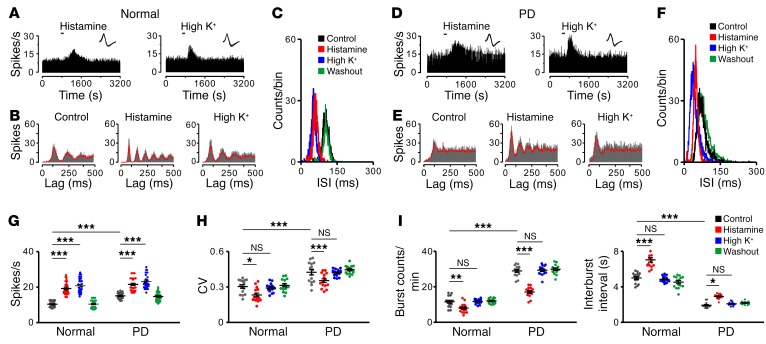
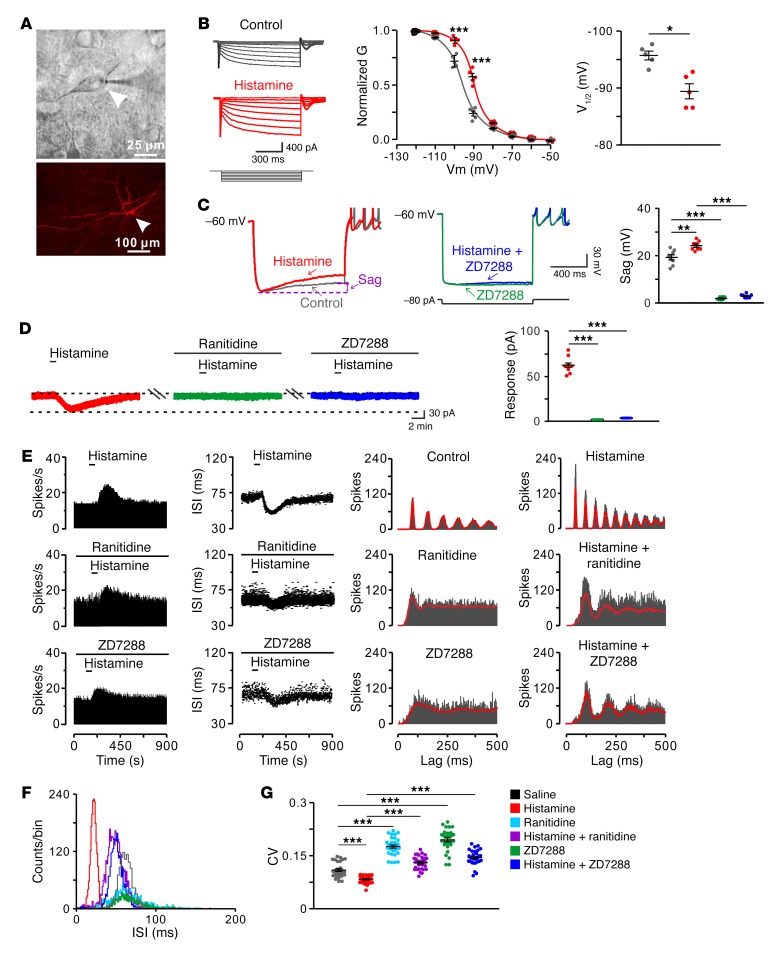
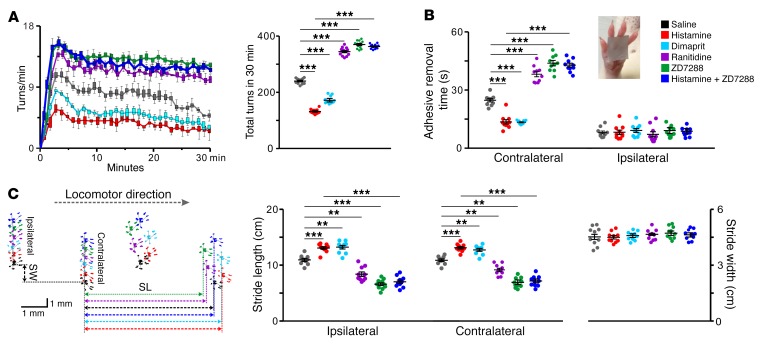
The subthalamic nucleus (STN) is an effective therapeutic target for deep brain stimulation (DBS) for Parkinson's disease (PD), and histamine levels are elevated in the basal ganglia in PD patients. However, the effect of endogenous histaminergic modulation on STN neuronal activities and the neuronal mechanism underlying STN-DBS are unknown. Here, we report that STN neuronal firing patterns are more crucial than firing rates for motor control. Histamine excited STN neurons, but paradoxically ameliorated parkinsonian motor deficits, which we attributed to regularizing firing patterns of STN neurons via the hyperpolarization-activated cyclic nucleotide-gated channel 2 (HCN2) channel coupled to the H2 receptor. Intriguingly, DBS increased histamine release in the STN and regularized STN neuronal firing patterns under parkinsonian conditions. HCN2 contributed to the DBS-induced regularization of neuronal firing patterns, suppression of excessive β oscillations, and alleviation of motor deficits in PD. The results reveal an indispensable role for regularizing STN neuronal firing patterns in amelioration of parkinsonian motor dysfunction and a functional compensation for histamine in parkinsonian basal ganglia circuitry. The findings provide insights into mechanisms of STN-DBS as well as potential therapeutic targets and STN-DBS strategies for PD.
Keywords: Neuroscience; Parkinson’s disease.
Conflict of interest statement
Figures
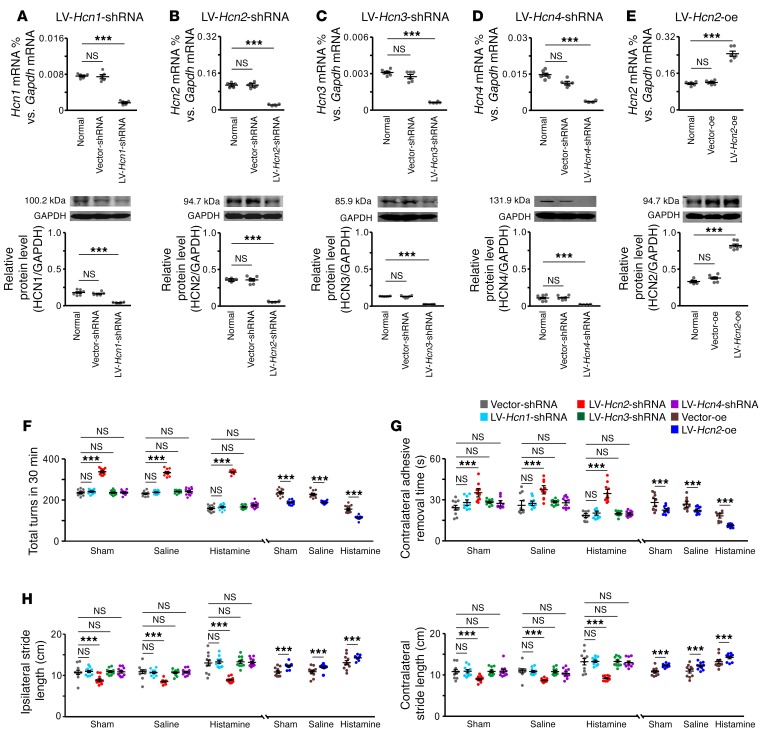
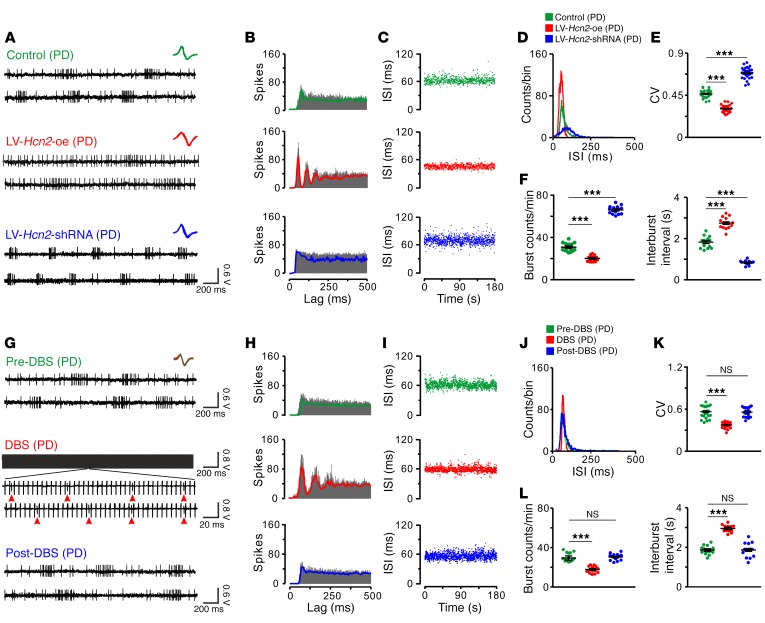
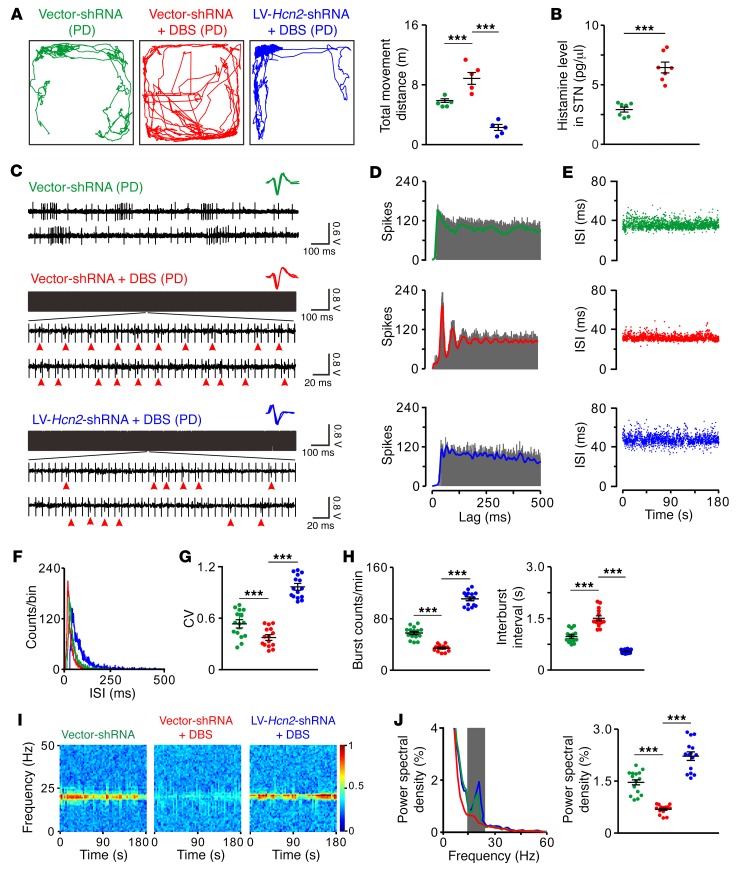
Comment in
-
Histamine and deep brain stimulation: the pharmacology of regularizing a brain.J Clin Invest. 2018 Dec 3;128(12):5201-5202. doi: 10.1172/JCI124777. Epub 2018 Oct 29. J Clin Invest. 2018. PMID: 30371507 Free PMC article.
Similar articles
-
Frequency-Specific Optogenetic Deep Brain Stimulation of Subthalamic Nucleus Improves Parkinsonian Motor Behaviors.J Neurosci. 2020 May 27;40(22):4323-4334. doi: 10.1523/JNEUROSCI.3071-19.2020. Epub 2020 Apr 20. J Neurosci. 2020. PMID: 32312888 Free PMC article.
-
Impaired glutamatergic projection from the motor cortex to the subthalamic nucleus in 6-hydroxydopamine-lesioned hemi-parkinsonian rats.Exp Neurol. 2018 Feb;300:135-148. doi: 10.1016/j.expneurol.2017.11.006. Epub 2017 Nov 8. Exp Neurol. 2018. PMID: 29126889
-
High-Frequency Stimulation of the Subthalamic Nucleus Activates Motor Cortex Pyramidal Tract Neurons by a Process Involving Local Glutamate, GABA and Dopamine Receptors in Hemi-Parkinsonian Rats.Chin J Physiol. 2018 Apr 30;61(2):92-105. doi: 10.4077/CJP.2018.BAG561. Chin J Physiol. 2018. PMID: 29590745
-
Subthalamic burst firing: A pathophysiological target in Parkinson's disease.Neurosci Biobehav Rev. 2022 Jan;132:410-419. doi: 10.1016/j.neubiorev.2021.11.044. Epub 2021 Nov 29. Neurosci Biobehav Rev. 2022. PMID: 34856222 Review.
-
[Subthalamic nucleus: from circuits, functions to a deep brain stimulation target for the treatment of Parkinson's disease].Sheng Li Xue Bao. 2017 Oct 25;69(5):611-622. Sheng Li Xue Bao. 2017. PMID: 29063109 Review. Chinese.
Cited by
-
Electroceutically induced subthalamic high-frequency oscillations and evoked compound activity may explain the mechanism of therapeutic stimulation in Parkinson's disease.Commun Biol. 2021 Mar 23;4(1):393. doi: 10.1038/s42003-021-01915-7. Commun Biol. 2021. PMID: 33758361 Free PMC article.
-
Nucleus accumbens ghrelin signaling controls anxiety-like behavioral response to acute stress.Behav Brain Funct. 2024 Jul 4;20(1):18. doi: 10.1186/s12993-024-00244-z. Behav Brain Funct. 2024. PMID: 38965529 Free PMC article.
-
Maladaptive Downregulation of Autonomous Subthalamic Nucleus Activity following the Loss of Midbrain Dopamine Neurons.Cell Rep. 2019 Jul 23;28(4):992-1002.e4. doi: 10.1016/j.celrep.2019.06.076. Cell Rep. 2019. PMID: 31340159 Free PMC article.
-
Differential modulation of subthalamic projection neurons by short-term and long-term electrical stimulation in physiological and parkinsonian conditions.Acta Pharmacol Sin. 2022 Aug;43(8):1928-1939. doi: 10.1038/s41401-021-00811-4. Epub 2021 Dec 8. Acta Pharmacol Sin. 2022. PMID: 34880404 Free PMC article.
-
Role of HCN channels in the functions of basal ganglia and Parkinson's disease.Cell Mol Life Sci. 2024 Mar 13;81(1):135. doi: 10.1007/s00018-024-05163-w. Cell Mol Life Sci. 2024. PMID: 38478096 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
