

ATRX loss induces multiple hallmarks of the alternative lengthening of telomeres (ALT) phenotype in human glioma cell lines in a cell line-specific manner
- PMID: 30226859
- PMCID: PMC6143253
- DOI: 10.1371/journal.pone.0204159
ATRX loss induces multiple hallmarks of the alternative lengthening of telomeres (ALT) phenotype in human glioma cell lines in a cell line-specific manner
Abstract
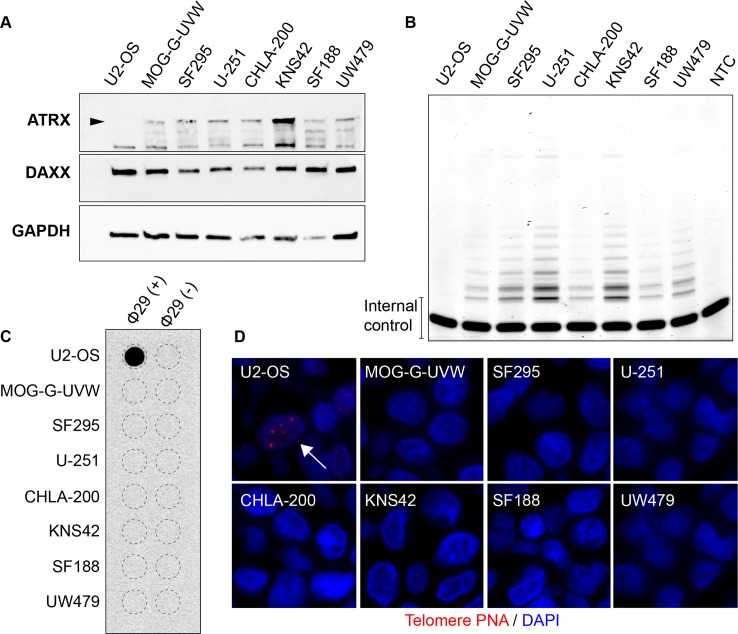
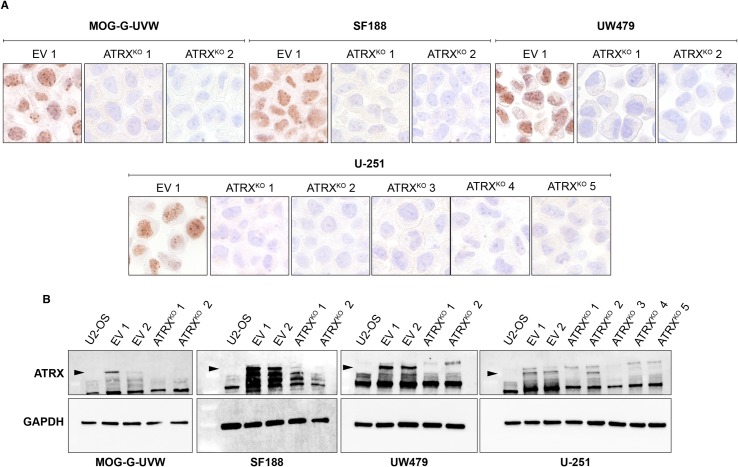
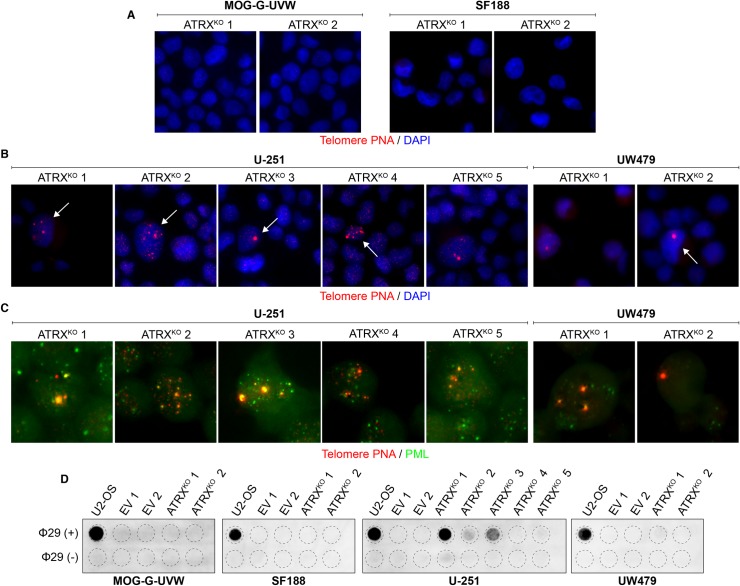
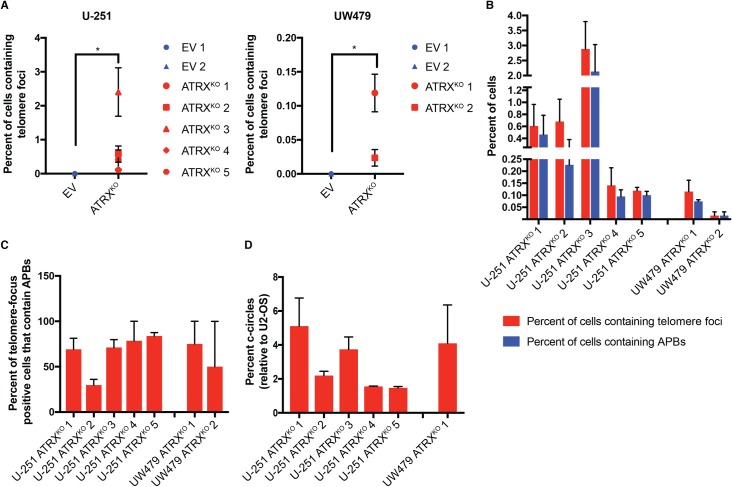
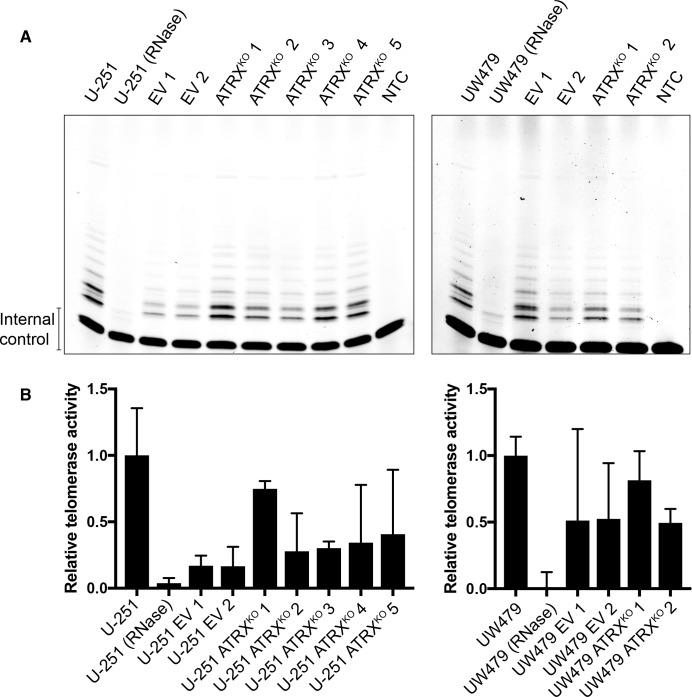
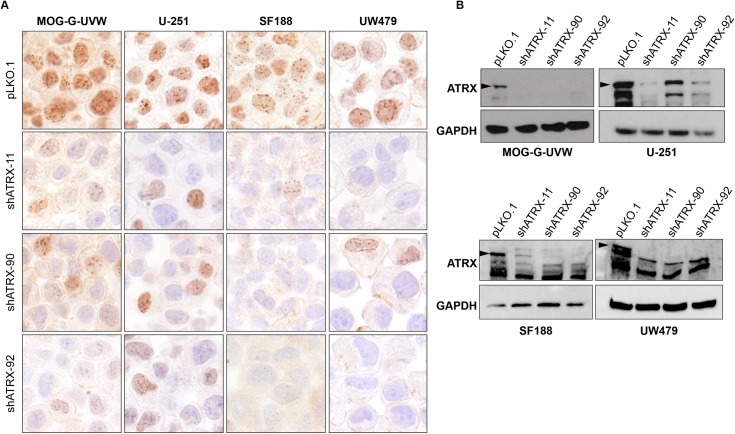
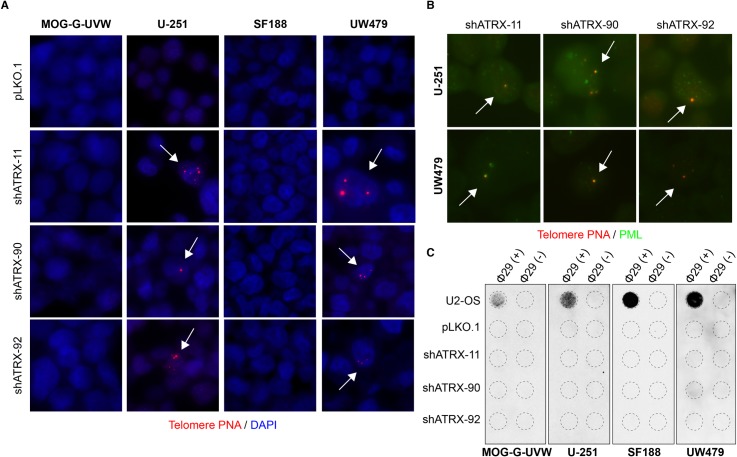
Cancers must maintain their telomeres at lengths sufficient for cell survival. In several cancer subtypes, a recombination-like mechanism termed alternative lengthening of telomeres (ALT), is frequently used for telomere length maintenance. Cancers utilizing ALT often have lost functional ATRX, a chromatin remodeling protein, through mutation or deletion, thereby strongly implicating ATRX as an ALT suppressor. Herein, we have generated functional ATRX knockouts in four telomerase-positive, ALT-negative human glioma cell lines: MOG-G-UVW, SF188, U-251 and UW479. After loss of ATRX, two of the four cell lines (U-251 and UW479) show multiple characteristics of ALT-positive cells, including ultrabright telomeric DNA foci, ALT-associated PML bodies, and c-circles. However, telomerase activity and overall telomere length heterogeneity are unaffected after ATRX loss, regardless of cellular context. The two cell lines that showed ALT hallmarks after complete ATRX loss also did so upon ATRX depletion via shRNA-mediated knockdown. These results suggest that other genomic or epigenetic events, in addition to ATRX loss, are necessary for the induction of ALT in human cancer.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Hayflick L. The limited in vitro lifetime of human diploid cell strains. Exp Cell Res. 1965;37:614–36. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
