

The piRNA pathway responds to environmental signals to establish intergenerational adaptation to stress
- PMID: 30227863
- PMCID: PMC6145337
- DOI: 10.1186/s12915-018-0571-y
The piRNA pathway responds to environmental signals to establish intergenerational adaptation to stress
Abstract
Background: piRNAs have a constitutive role in genome defence by silencing transposable elements in the germline. In the nematode Caenorhabditis elegans, piRNAs also induce epigenetic silencing of transgenes, which can be maintained for many generations in the absence of the piRNA pathway. The role of multi-generational epigenetic inheritance in adaptation to the environment is unknown.
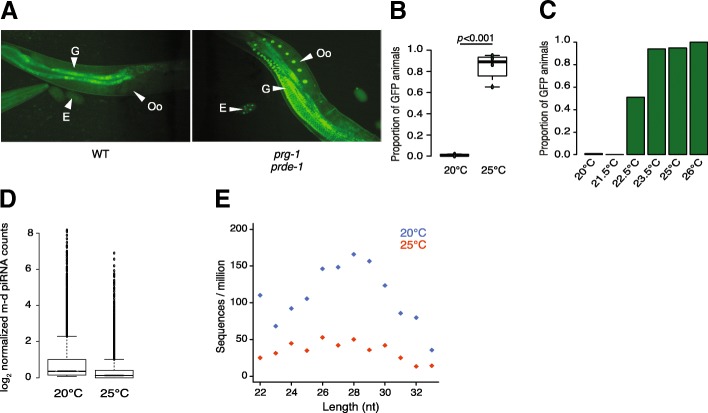
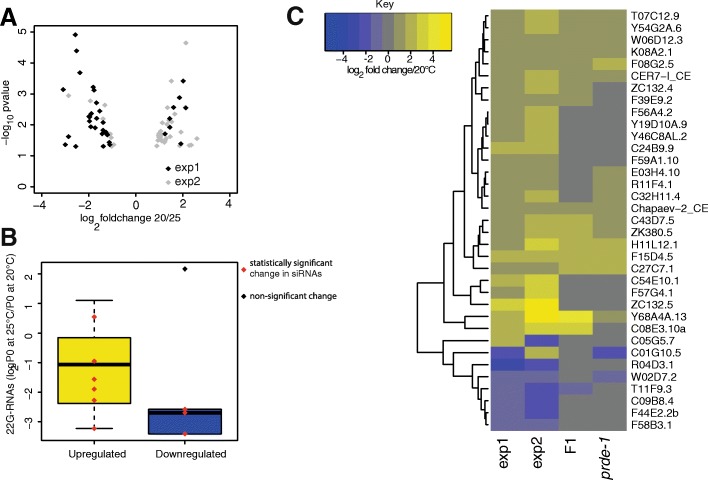
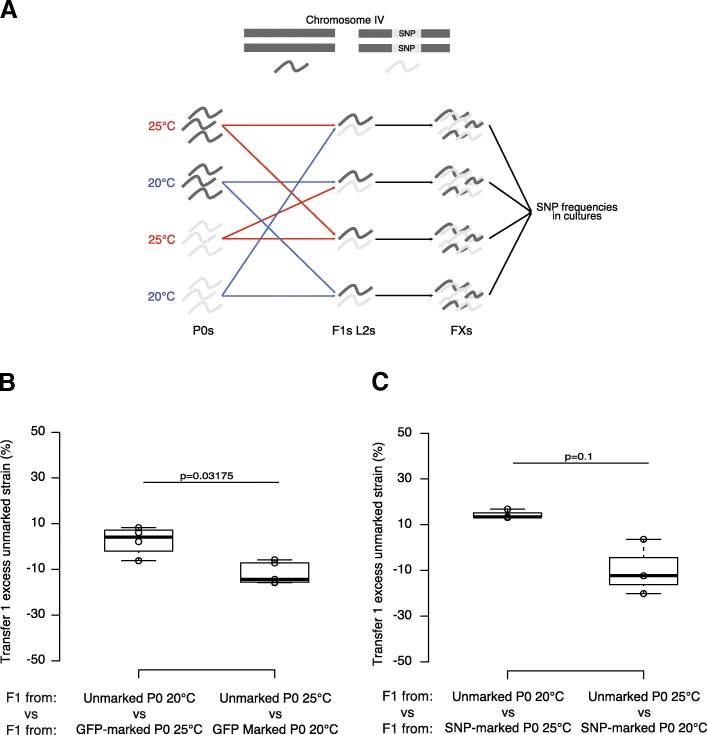
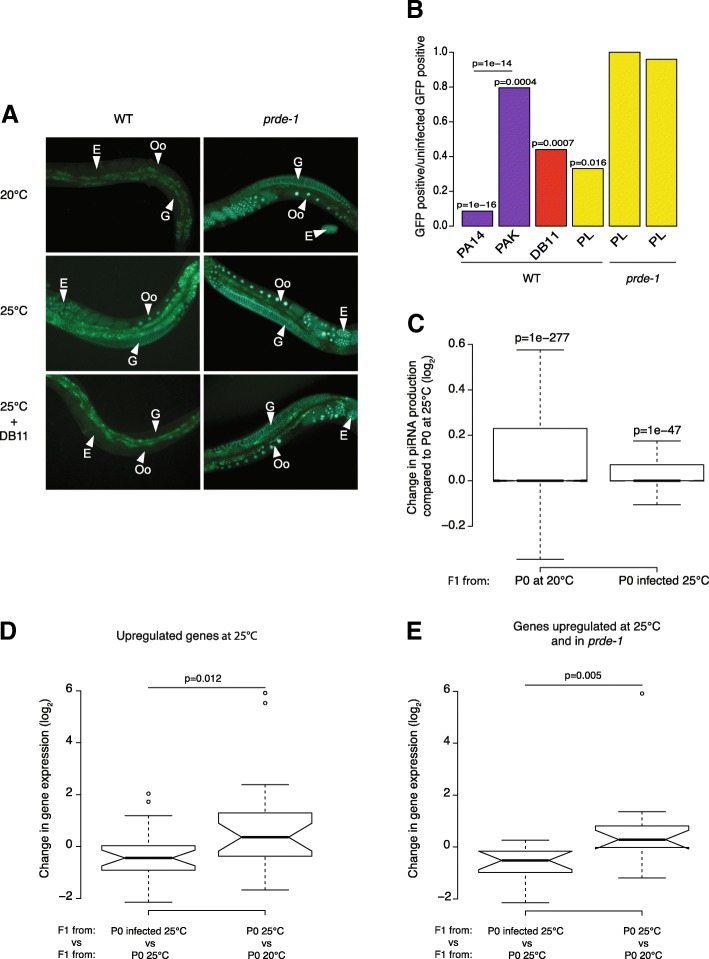
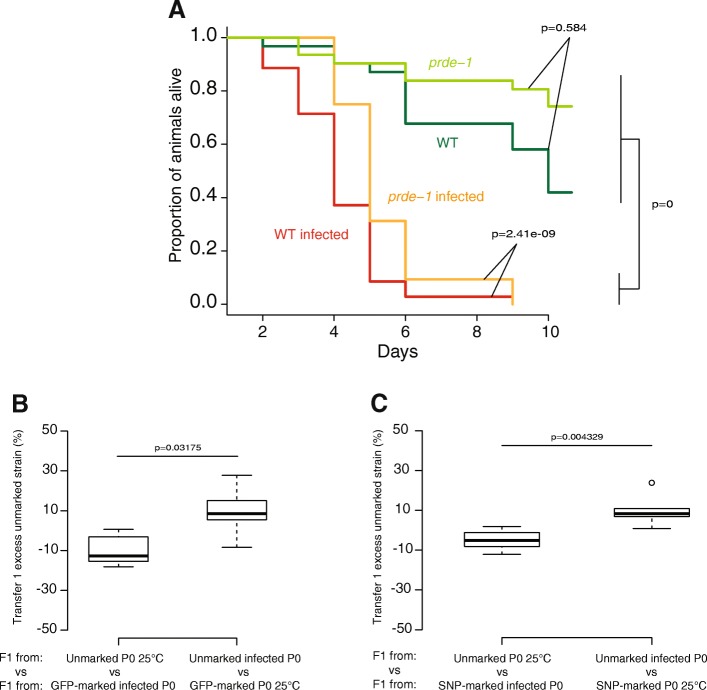
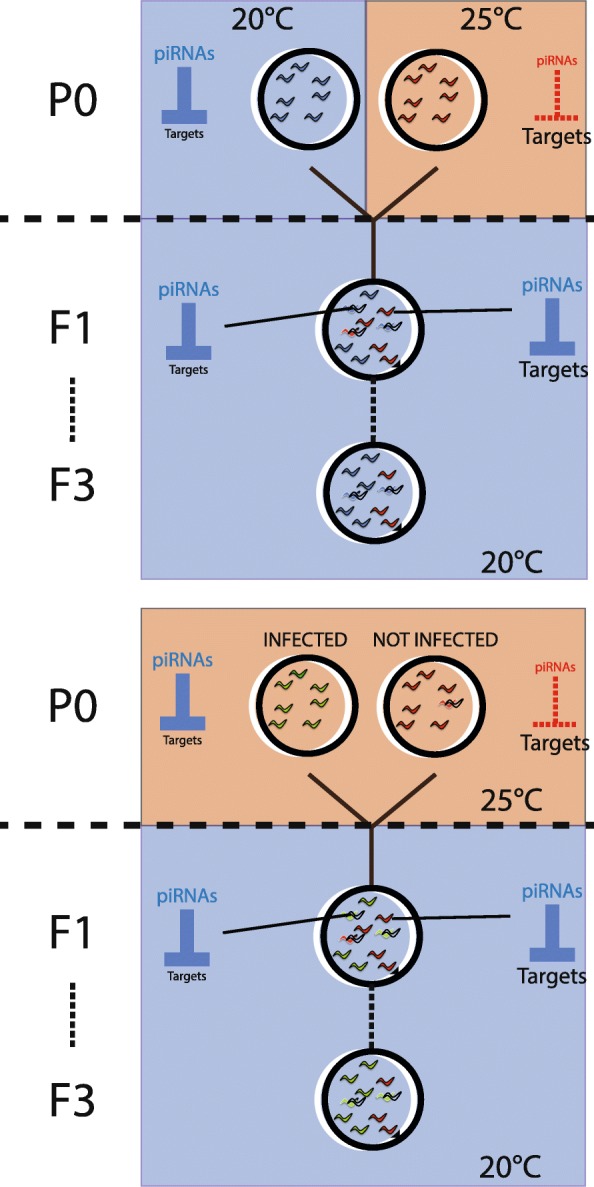
Results: Here, we show that piRNA biogenesis is downregulated in response to a small increase in temperature. Some effects on gene expression persist into subsequent generations and are associated with a negative fitness cost. We show that simultaneous infection with pathogenic bacteria suppresses downregulation of the piRNA pathway in response to increased temperature. This effect is associated with increased fitness of progeny of infected animals in subsequent generations.
Conclusions: Our results show that the piRNA pathway integrates inputs from the environment to establish intergenerational responses to environmental conditions, with important consequences for the fitness of the subsequent generation.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Eggert H., Kurtz J., Diddens-de Buhr M. F. Different effects of paternal trans-generational immune priming on survival and immunity in step and genetic offspring. Proceedings of the Royal Society B: Biological Sciences. 2014;281(1797):20142089–20142089. doi: 10.1098/rspb.2014.2089. - DOI - PMC - PubMed
-
- Radford E. J., Ito M., Shi H., Corish J. A., Yamazawa K., Isganaitis E., Seisenberger S., Hore T. A., Reik W., Erkek S., Peters A. H. F. M., Patti M.-E., Ferguson-Smith A. C. In utero undernourishment perturbs the adult sperm methylome and intergenerational metabolism. Science. 2014;345(6198):1255903–1255903. doi: 10.1126/science.1255903. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
