

Extracellular Phosphorylation of TIMP-2 by Secreted c-Src Tyrosine Kinase Controls MMP-2 Activity
- PMID: 30227959
- PMCID: PMC6135941
- DOI: 10.1016/j.isci.2018.02.004
Extracellular Phosphorylation of TIMP-2 by Secreted c-Src Tyrosine Kinase Controls MMP-2 Activity
Abstract
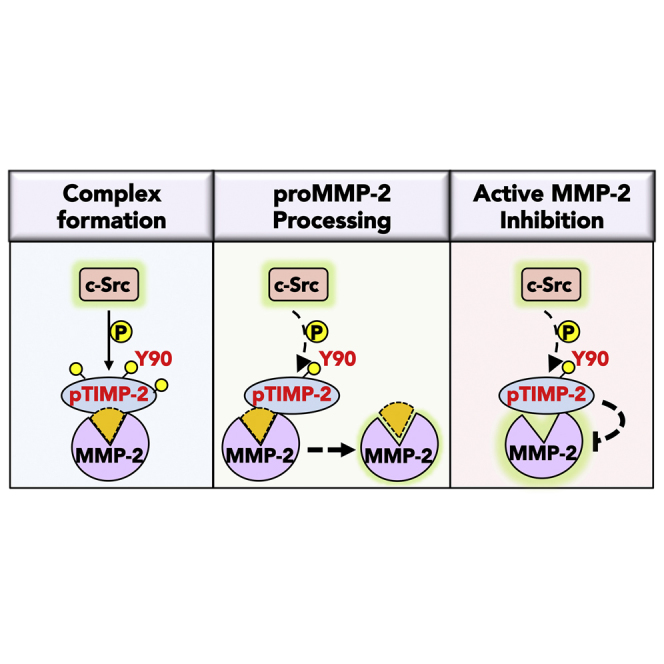
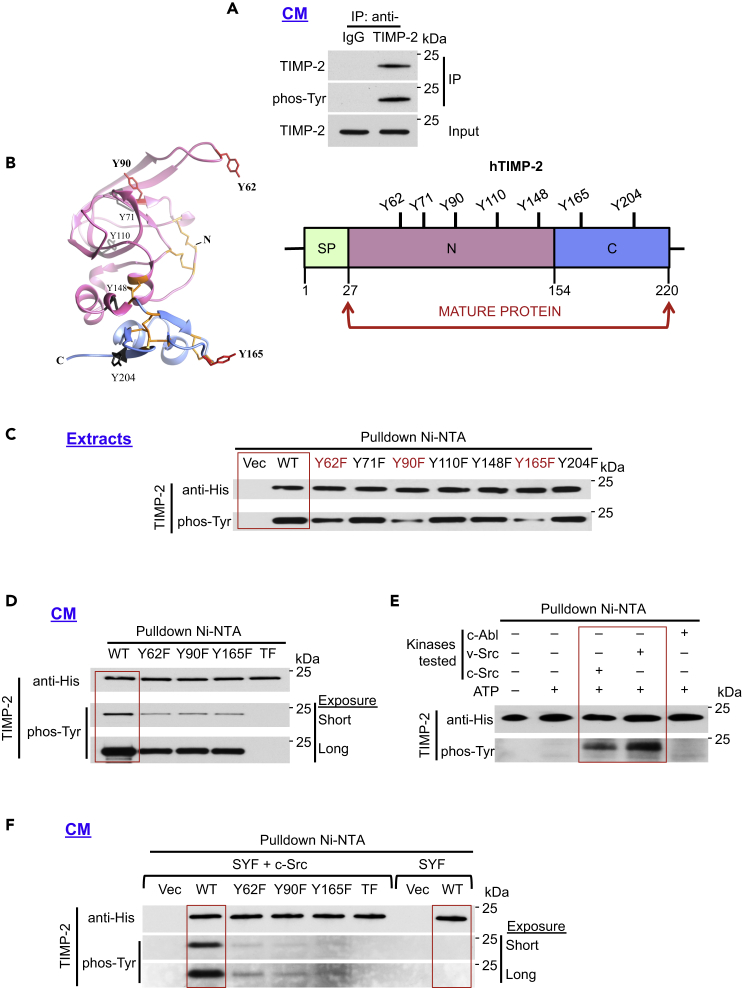
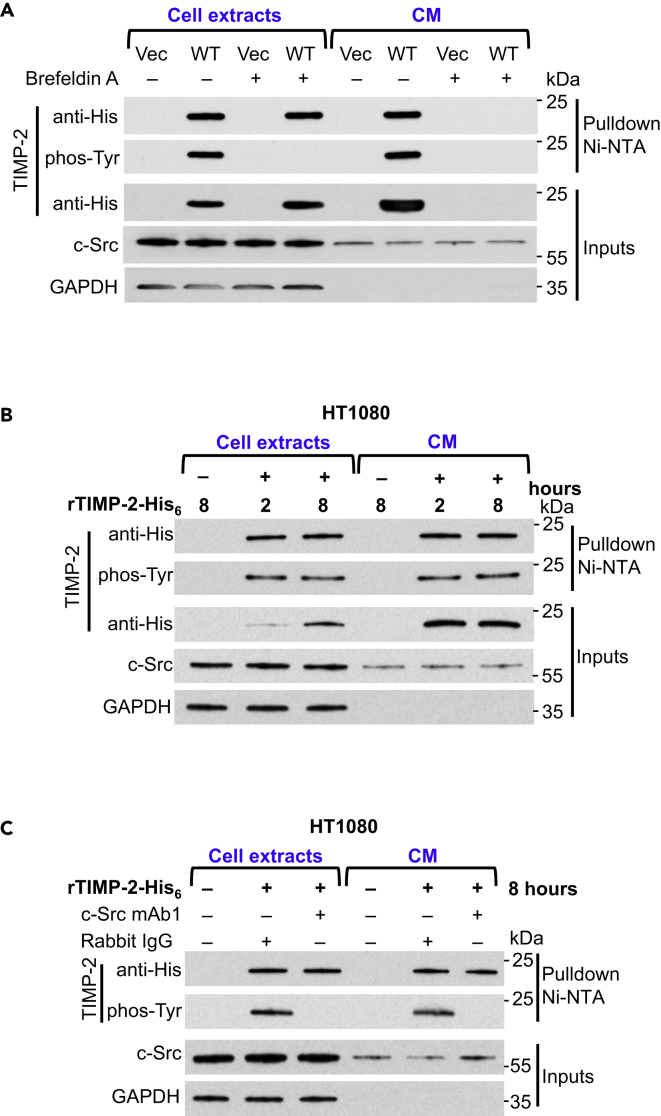
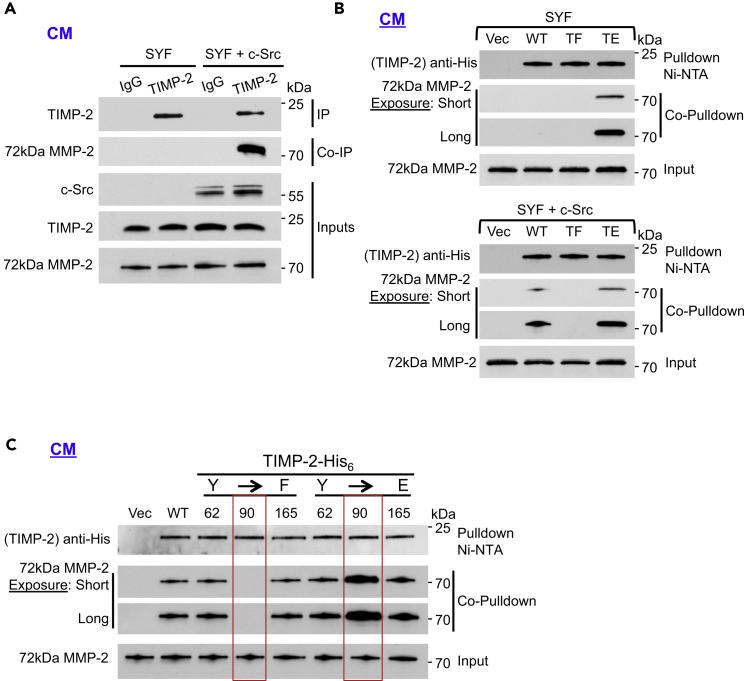
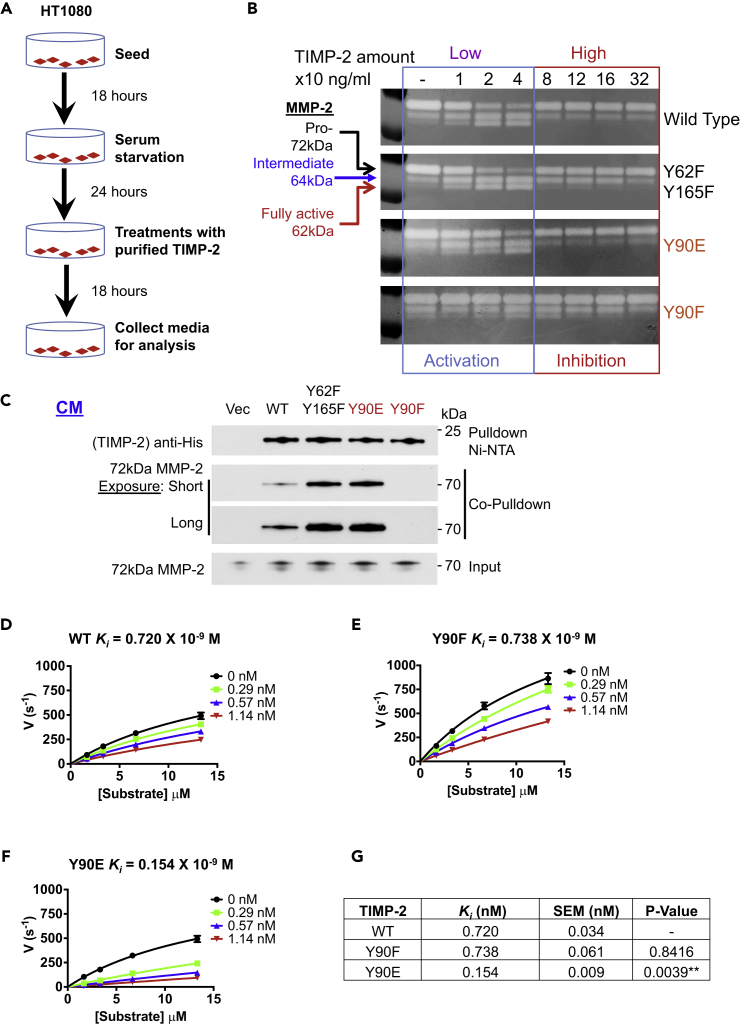
The tissue inhibitor of metalloproteinases 2 (TIMP-2) is a specific endogenous inhibitor of matrix metalloproteinase 2 (MMP-2), which is a key enzyme that degrades the extracellular matrix and promotes tumor cell invasion. Although the TIMP-2:MMP-2 complex controls proteolysis, the signaling mechanism by which the two proteins associate in the extracellular space remains unidentified. Here we report that TIMP-2 is phosphorylated outside the cell by secreted c-Src tyrosine kinase. As a consequence, phosphorylation at Y90 significantly enhances TIMP-2 potency as an MMP-2 inhibitor and weakens the catalytic action of the active enzyme. TIMP-2 phosphorylation also appears to be essential for its interaction with the latent enzyme proMMP-2 in vivo. Absence of the kinase or non-phosphorylatable Y90 abolishes TIMP-2 binding to the latent enzyme, ultimately hampering proMMP-2 activation. Together, TIMP-2 phosphorylation by secreted c-Src represents a critical extracellular regulatory mechanism that controls the proteolytic function of MMP-2.
Keywords: Biochemistry; Enzymology; Molecular Biology.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Beebe K., Mollapour M., Scroggins B., Prodromou C., Xu W., Tokita M., Taldone T., Pullen L., Zierer B.K., Lee M.J. Posttranslational modification and conformational state of heat shock protein 90 differentially affect binding of chemically diverse small molecule inhibitors. Oncotarget. 2013;4:1065–1074. - PMC - PubMed
-
- Betts M.J., Russell R.B. Amino acid properties and consequences of substitutions. In: Barnes M.R., Gray I.C., editors. Bioinformatics for Geneticists. John Wiley & Sons, Ltd; 2003. pp. 291–316.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
