

The C-terminal GGAP motif of Hsp70 mediates substrate recognition and stress response in yeast
- PMID: 30228181
- PMCID: PMC6240856
- DOI: 10.1074/jbc.RA118.002691
The C-terminal GGAP motif of Hsp70 mediates substrate recognition and stress response in yeast
Abstract
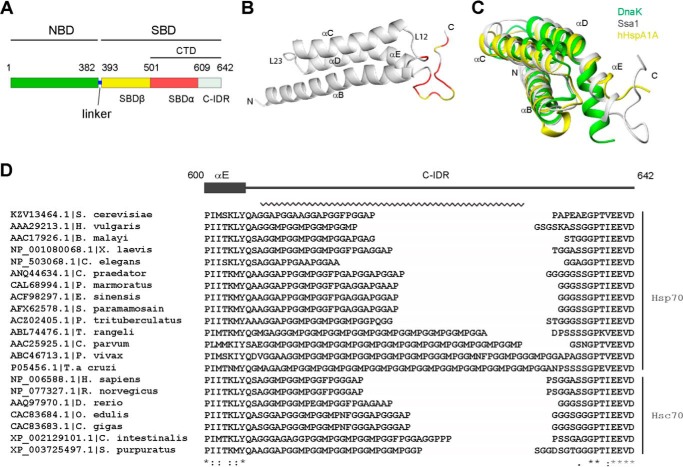
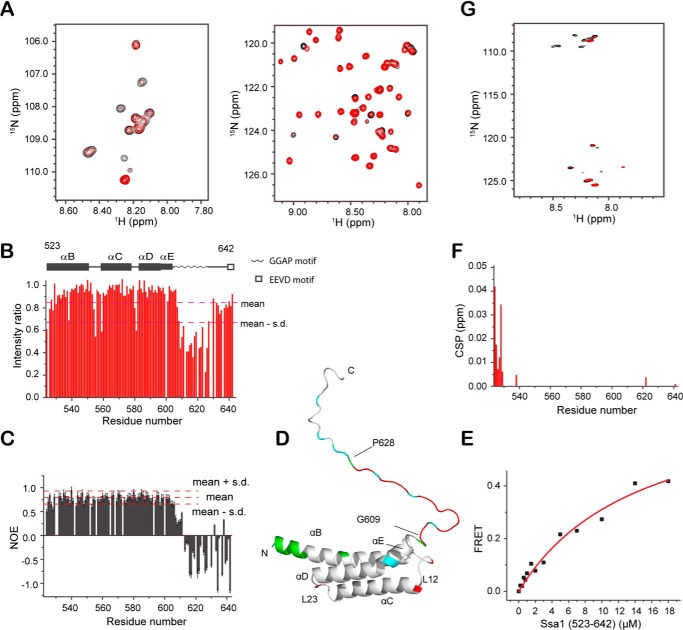
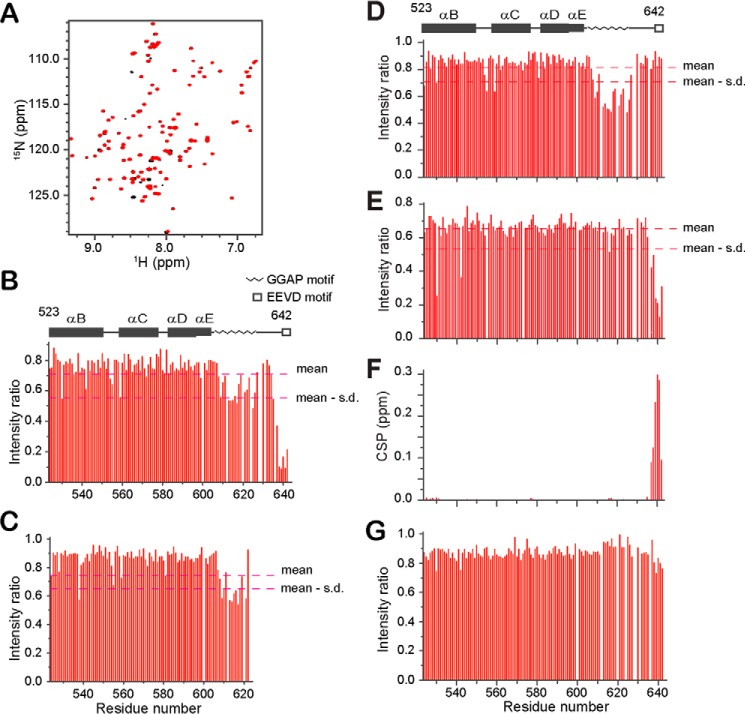
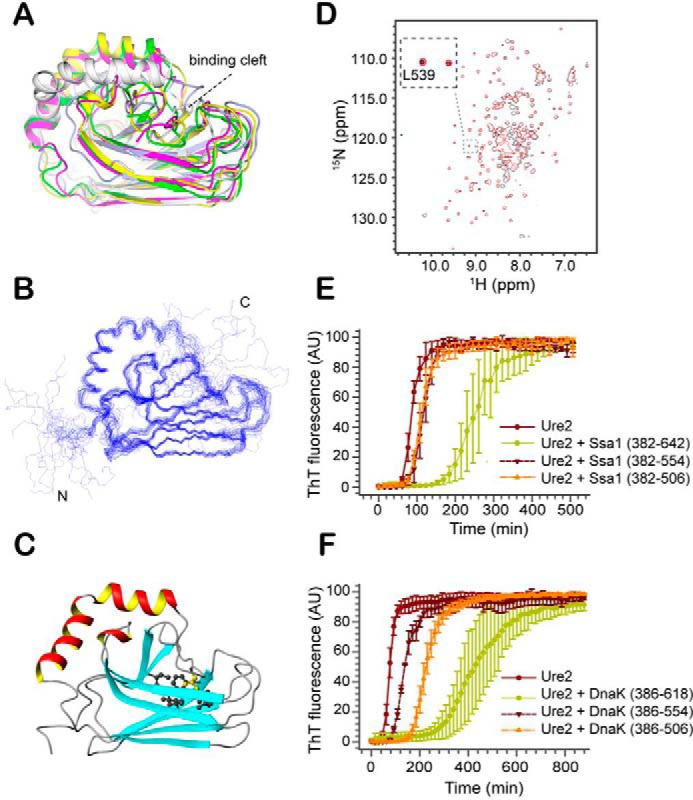
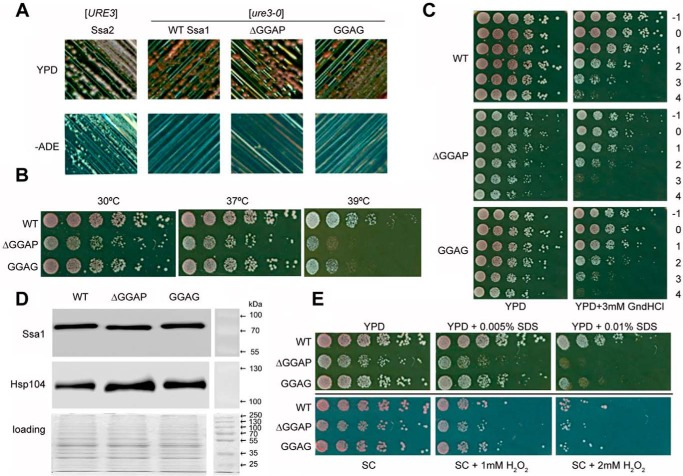
The allosteric coupling of the highly conserved nucleotide- and substrate-binding domains of Hsp70 has been studied intensively. In contrast, the role of the disordered, highly variable C-terminal region of Hsp70 remains unclear. In many eukaryotic Hsp70s, the extreme C-terminal EEVD motif binds to the tetratricopeptide-repeat domains of Hsp70 co-chaperones. Here, we discovered that the TVEEVD sequence of Saccharomyces cerevisiae cytoplasmic Hsp70 (Ssa1) functions as a SUMO-interacting motif. A second C-terminal motif of ∼15 amino acids between the α-helical lid and the extreme C terminus, previously identified in bacterial and eukaryotic organellar Hsp70s, is known to enhance chaperone function by transiently interacting with folding clients. Using structural analysis, interaction studies, fibril formation assays, and in vivo functional assays, we investigated the individual contributions of the α-helical bundle and the C-terminal disordered region of Ssa1 in the inhibition of fibril formation of the prion protein Ure2. Our results revealed that although the α-helical bundle of the Ssa1 substrate-binding domain (SBDα) does not directly bind to Ure2, the SBDα enhances the ability of Hsp70 to inhibit fibril formation. We found that a 20-residue C-terminal motif in Ssa1, containing GGAP and GGAP-like tetrapeptide repeats, can directly bind to Ure2, the Hsp40 co-chaperone Ydj1, and α-synuclein, but not to the SUMO-like protein SMT3 or BSA. Deletion or substitution of the Ssa1 GGAP motif impaired yeast cell tolerance to temperature and cell-wall damage stress. This study highlights that the C-terminal GGAP motif of Hsp70 is important for substrate recognition and mediation of the heat shock response.
Keywords: SUMO-interacting motif (SIM); Saccharomyces cerevisiae; amyloid; chaperone; heat shock protein (HSP); nuclear magnetic resonance (NMR); prion; proline-rich motif; protein structure; stress response.
© 2018 Gong et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases