

Structure-Stability-Function Mechanistic Links in the Anti-Measles Virus Action of Tocopherol-Derivatized Peptide Nanoparticles
- PMID: 30230818
- PMCID: PMC6399014
- DOI: 10.1021/acsnano.8b01422
Structure-Stability-Function Mechanistic Links in the Anti-Measles Virus Action of Tocopherol-Derivatized Peptide Nanoparticles
Abstract
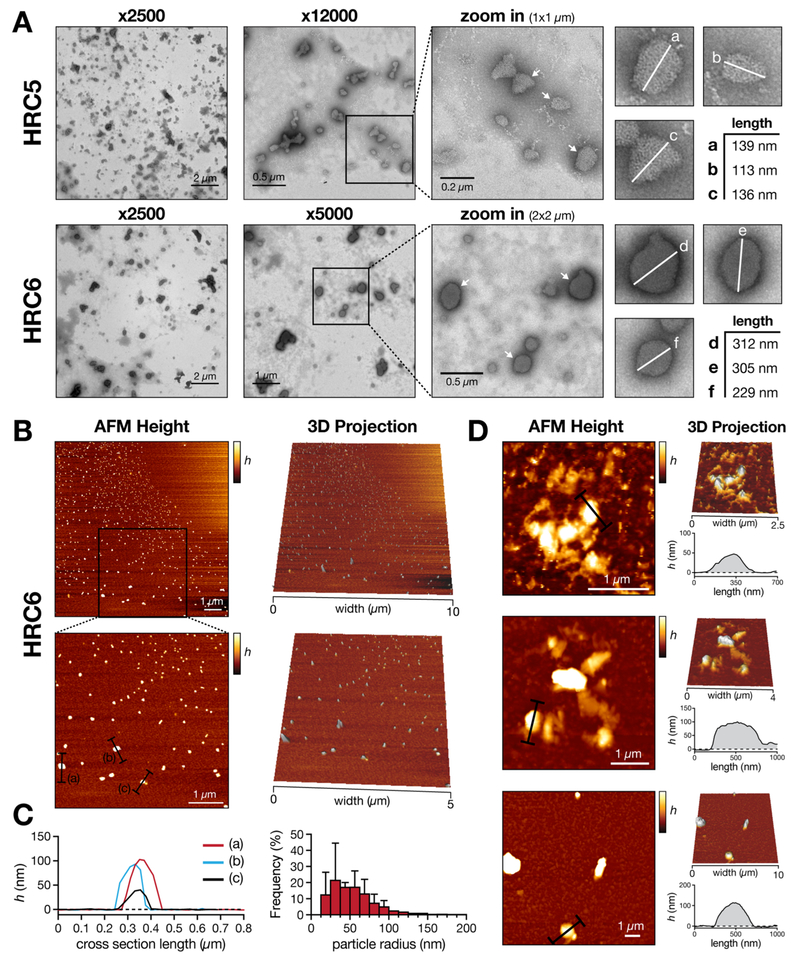
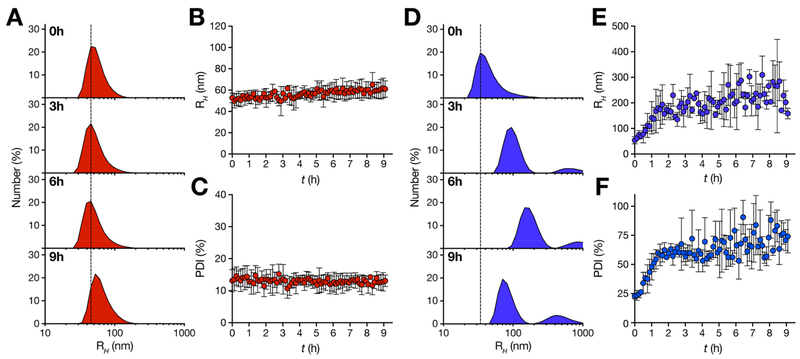
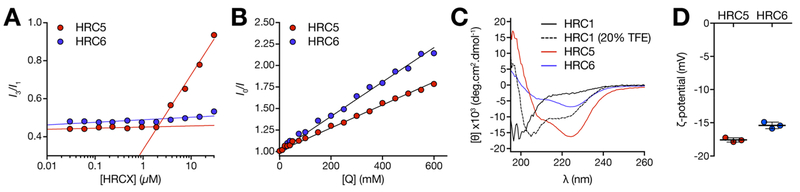
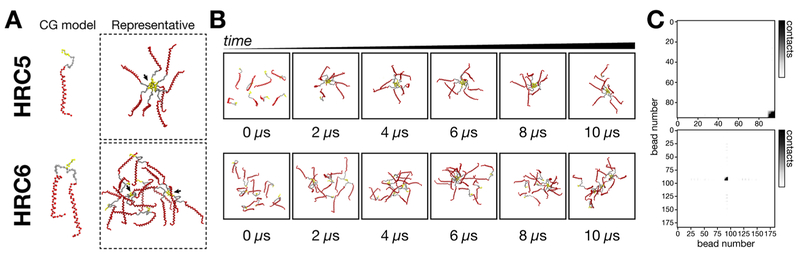
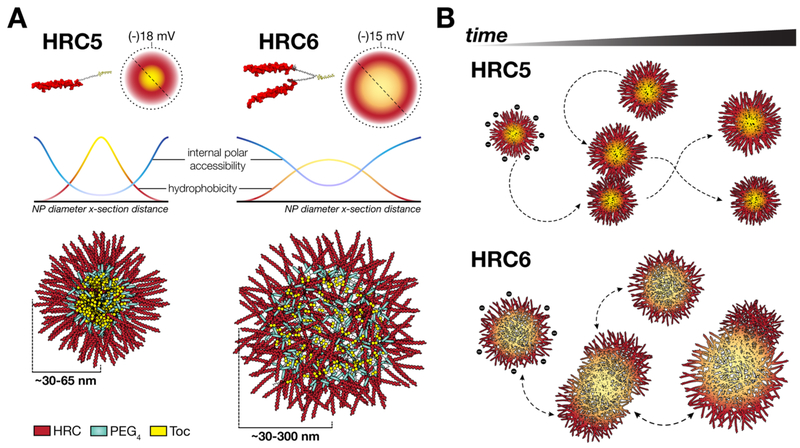
Measles remains one of the leading causes of child mortality worldwide and is re-emerging in some countries due to poor vaccine coverage, concomitant with importation of measles virus (MV) from endemic areas. The lack of specific chemotherapy contributes to negative outcomes, especially in infants or immunodeficient individuals. Fusion inhibitor peptides derived from the MV Fusion protein C-terminal Heptad Repeat (HRC) targeting MV envelope fusion glycoproteins block infection at the stage of entry into host cells, thus preventing viral multiplication. To improve efficacy of such entry inhibitors, we have modified a HRC peptide inhibitor by introducing properties of self-assembly into nanoparticles (NP) and higher affinity for both viral and cell membranes. Modification of the peptide consisted of covalent grafting with tocopherol to increase amphipathicity and lipophilicity (HRC5). One additional peptide inhibitor consisting of a peptide dimer grafted to tocopherol was also used (HRC6). Spectroscopic, imaging, and simulation techniques were used to characterize the NP and explore the molecular basis for their antiviral efficacy. HRC5 forms micellar stable NP while HRC6 aggregates into amorphous, loose, unstable NP. Interpeptide cluster bridging governs NP assembly into dynamic metastable states. The results are consistent with the conclusion that the improved efficacy of HRC6 relative to HRC5 can be attributed to NP instability, which leads to more extensive partition to target membranes and binding to viral target proteins.
Keywords: antiviral; fusion inhibitor; measles virus; metastable; nanoparticle; peptide; self-assembling.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
In Vivo Efficacy of Measles Virus Fusion Protein-Derived Peptides Is Modulated by the Properties of Self-Assembly and Membrane Residence.J Virol. 2016 Dec 16;91(1):e01554-16. doi: 10.1128/JVI.01554-16. Print 2017 Jan 1. J Virol. 2016. PMID: 27733647 Free PMC article.
-
Biophysical Properties and Antiviral Activities of Measles Fusion Protein Derived Peptide Conjugated with 25-Hydroxycholesterol.Molecules. 2017 Oct 31;22(11):1869. doi: 10.3390/molecules22111869. Molecules. 2017. PMID: 29088094 Free PMC article.
-
Prevention of measles virus infection by intranasal delivery of fusion inhibitor peptides.J Virol. 2015 Jan 15;89(2):1143-55. doi: 10.1128/JVI.02417-14. Epub 2014 Nov 5. J Virol. 2015. PMID: 25378493 Free PMC article.
-
Measles Virus Fusion Protein: Structure, Function and Inhibition.Viruses. 2016 Apr 21;8(4):112. doi: 10.3390/v8040112. Viruses. 2016. PMID: 27110811 Free PMC article. Review.
-
Prospects and strategies for the discovery and development of small-molecule inhibitors of six-helix bundle formation in class 1 viral fusion proteins.Curr Opin Investig Drugs. 2006 Feb;7(2):118-27. Curr Opin Investig Drugs. 2006. PMID: 16499281 Review.
Cited by
-
Carbosilane Dendritic Amphiphiles from Cholesterol or Vitamin E for Micelle Formation.Pharmaceutics. 2024 Mar 25;16(4):451. doi: 10.3390/pharmaceutics16040451. Pharmaceutics. 2024. PMID: 38675112 Free PMC article.
-
SARS-CoV-2 Immuno-Pathogenesis and Potential for Diverse Vaccines and Therapies: Opportunities and Challenges.Infect Dis Rep. 2021 Feb 4;13(1):102-125. doi: 10.3390/idr13010013. Infect Dis Rep. 2021. PMID: 33557330 Free PMC article. Review.
-
Intranasal fusion inhibitory lipopeptide prevents direct contact SARS-CoV-2 transmission in ferrets.bioRxiv [Preprint]. 2020 Nov 5:2020.11.04.361154. doi: 10.1101/2020.11.04.361154. bioRxiv. 2020. Update in: Science. 2021 Mar 26;371(6536):1379-1382. doi: 10.1126/science.abf4896. PMID: 33173865 Free PMC article. Updated. Preprint.
-
Enhancing the solubility of SARS-CoV-2 inhibitors to increase future prospects for clinical development.J Virol. 2025 Mar 18;99(3):e0215924. doi: 10.1128/jvi.02159-24. Epub 2025 Feb 4. J Virol. 2025. PMID: 39902960 Free PMC article.
-
Inhibition of Measles Viral Fusion Is Enhanced by Targeting Multiple Domains of the Fusion Protein.ACS Nano. 2021 Aug 24;15(8):12794-12803. doi: 10.1021/acsnano.1c02057. Epub 2021 Jul 22. ACS Nano. 2021. PMID: 34291895 Free PMC article.
References
-
- Moss WJ; Griffin DE Measles. Lancet 2012, 379, 153–164. - PubMed
-
- WHO. 2016 Midterm Review of the Global Vaccine Action Plan; 2016; pp 1–26.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
