

Molecular Characterization of the Viroporin Function of Foot-and-Mouth Disease Virus Nonstructural Protein 2B
- PMID: 30232178
- PMCID: PMC6232471
- DOI: 10.1128/JVI.01360-18
Molecular Characterization of the Viroporin Function of Foot-and-Mouth Disease Virus Nonstructural Protein 2B
Abstract
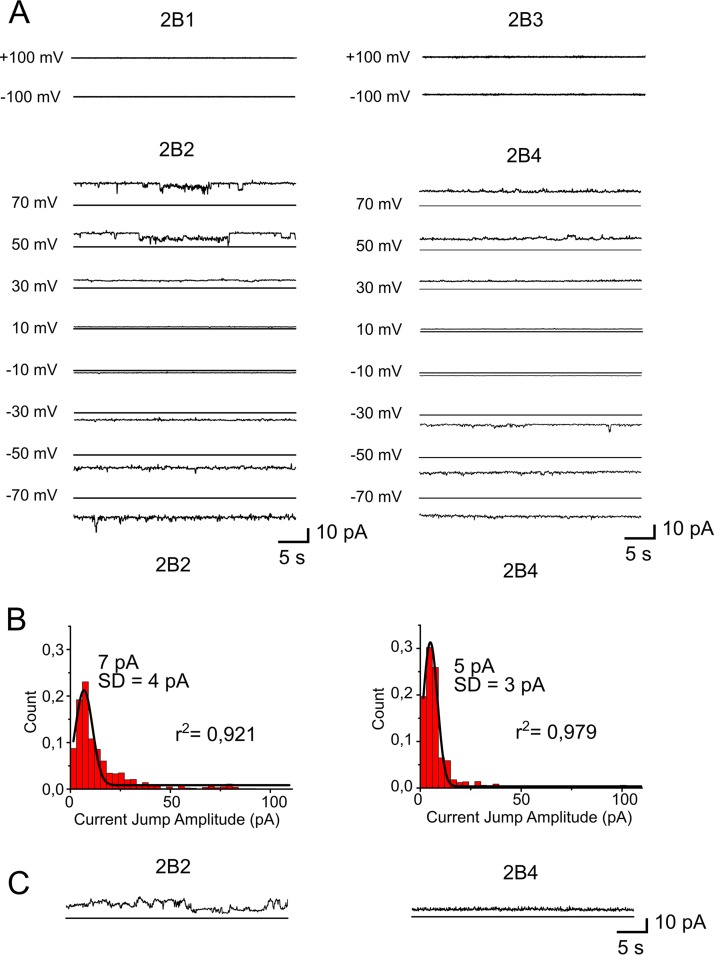
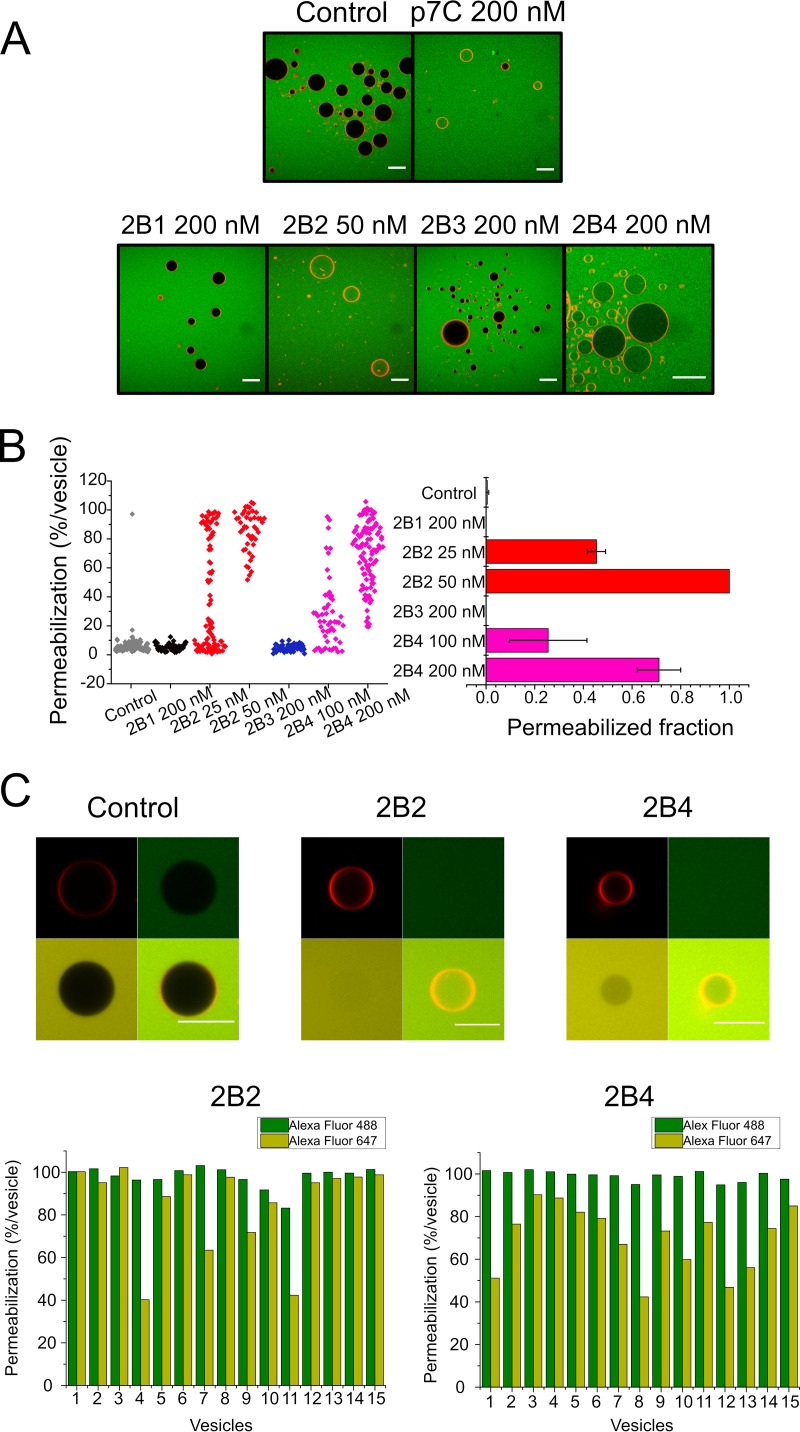
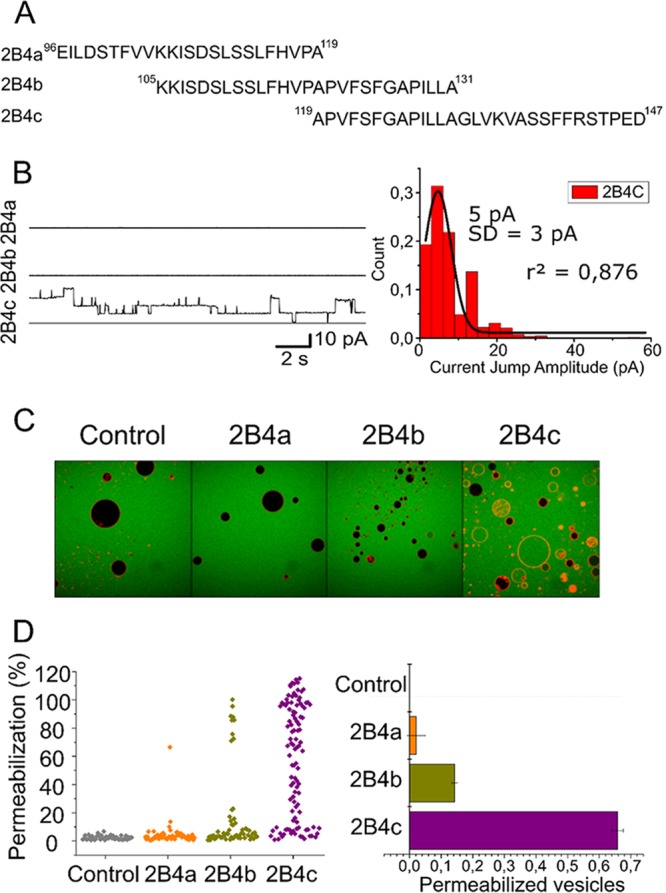
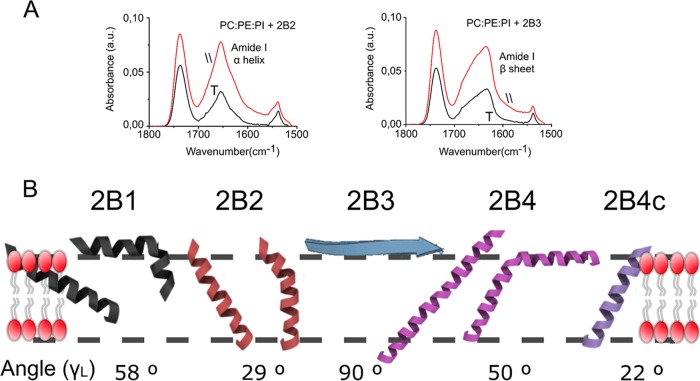
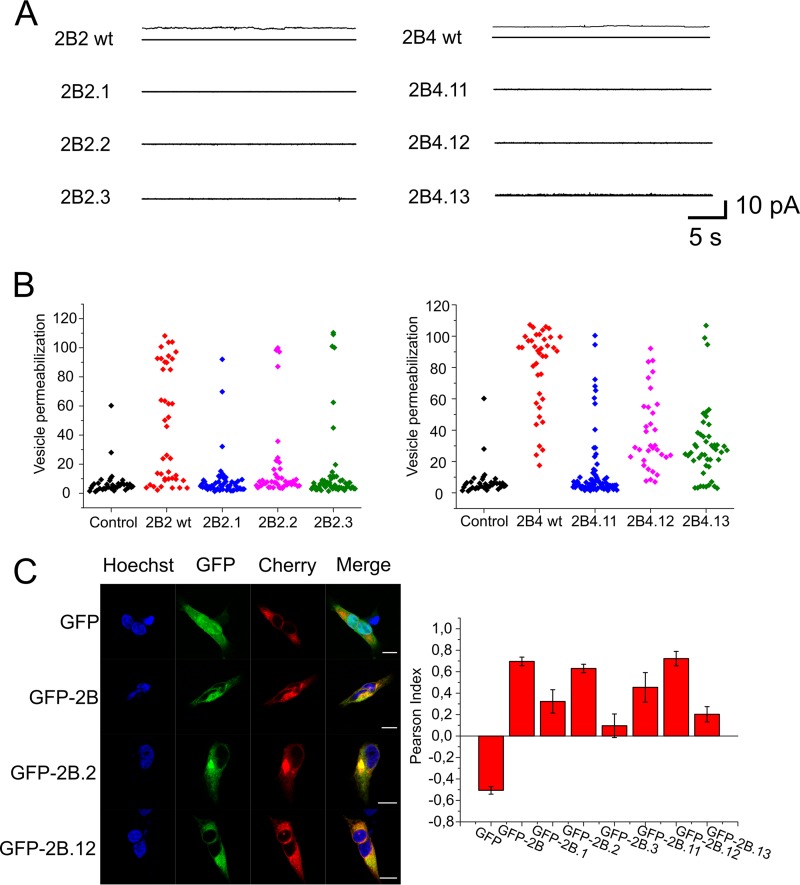
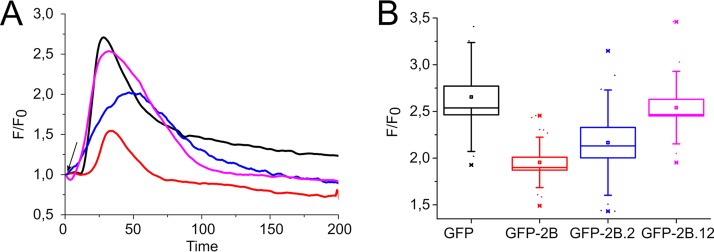
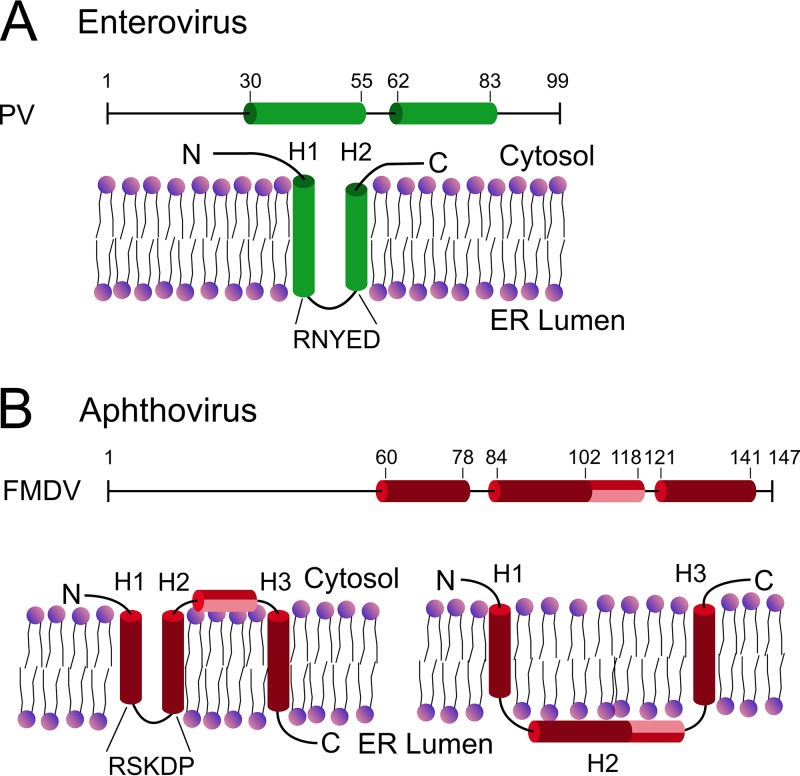
Nonstructural protein 2B of foot-and-mouth disease (FMD) virus (FMDV) is comprised of a small, hydrophobic, 154-amino-acid protein. Structure-function analyses demonstrated that FMDV 2B is an ion channel-forming protein. Infrared spectroscopy measurements using partially overlapping peptides that spanned regions between amino acids 28 and 147 demonstrated the adoption of helical conformations in two putative transmembrane regions between residues 60 and 78 and between residues 119 and 147 and a third transmembrane region between residues 79 and 106, adopting a mainly extended structure. Using synthetic peptides, ion channel activity measurements in planar lipid bilayers and imaging of single giant unilamellar vesicles (GUVs) revealed the existence of two sequences endowed with membrane-porating activity: one spanning FMDV 2B residues 55 to 82 and the other spanning the C-terminal region of 2B from residues 99 to 147. Mapping the latter sequence identified residues 119 to 147 as being responsible for the activity. Experiments to assess the degree of insertion of the synthetic peptides in bilayers and the inclination angle adopted by each peptide regarding the membrane plane normal confirm that residues 55 to 82 and 119 to 147 of 2B actively insert as transmembrane helices. Using reverse genetics, a panel of 13 FMD recombinant mutant viruses was designed, which harbored nonconservative as well as alanine substitutions in critical amino acid residues in the area between amino acid residues 28 and 147. Alterations to any of these structures interfered with pore channel activity and the capacity of the protein to permeabilize the endoplasmic reticulum (ER) to calcium and were lethal for virus replication. Thus, FMDV 2B emerges as the first member of the viroporin family containing two distinct pore domains.IMPORTANCE FMDV nonstructural protein 2B is able to insert itself into cellular membranes to form a pore. This pore allows the passage of ions and small molecules through the membrane. In this study, we were able to show that both current and small molecules are able to pass though the pore made by 2B. We also discovered for the first time a virus with a pore-forming protein that contains two independent functional pores. By making mutations in our infectious clone of FMDV, we determined that mutations in either pore resulted in nonviable virus. This suggests that both pore-forming functions are independently required during FMDV infection.
Keywords: 2B; FMD; FMDV; foot-and-mouth disease; viroporin.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- de Jong AS, Wessels E, Dijkman HB, Galama JM, Melchers WJ, Willems PH, van Kuppeveld FJ. 2003. Determinants for membrane association and permeabilization of the coxsackievirus 2B protein and the identification of the Golgi complex as the target organelle. J Biol Chem 278:1012–1021. doi:10.1074/jbc.M207745200. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
