

Necroptosis of infiltrated macrophages drives Yersinia pestis dispersal within buboes
- PMID: 30232285
- PMCID: PMC6237236
- DOI: 10.1172/jci.insight.122188
Necroptosis of infiltrated macrophages drives Yersinia pestis dispersal within buboes
Abstract
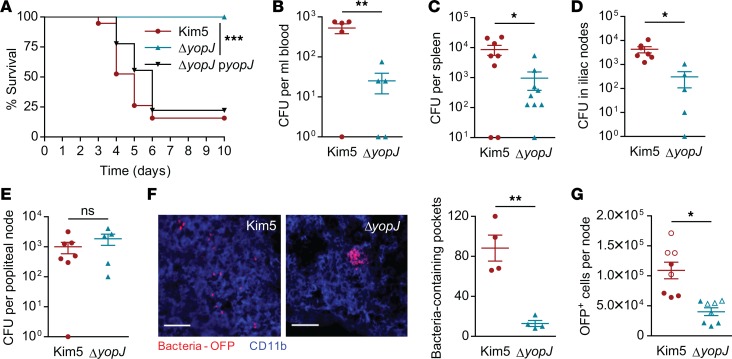
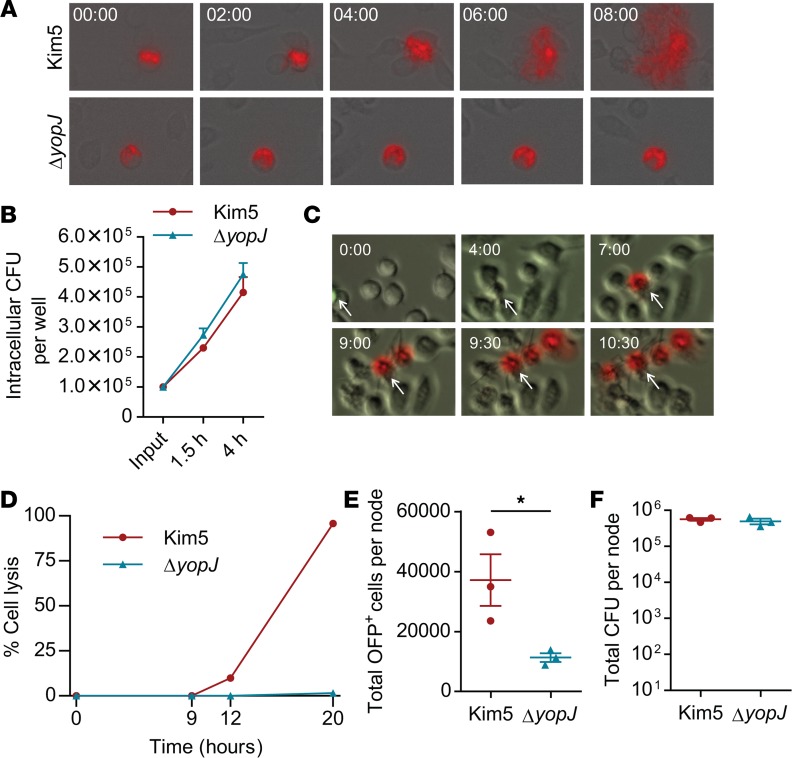
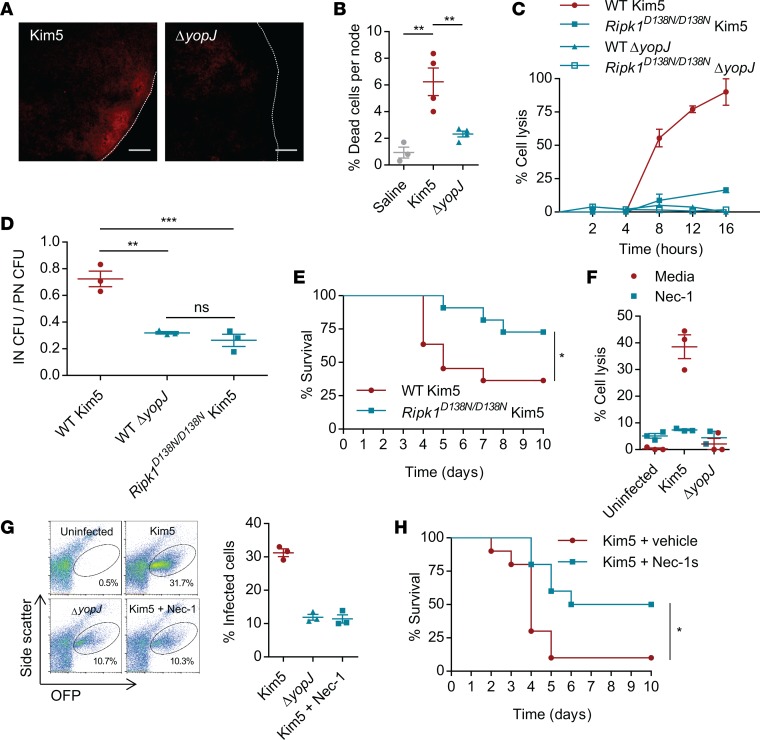
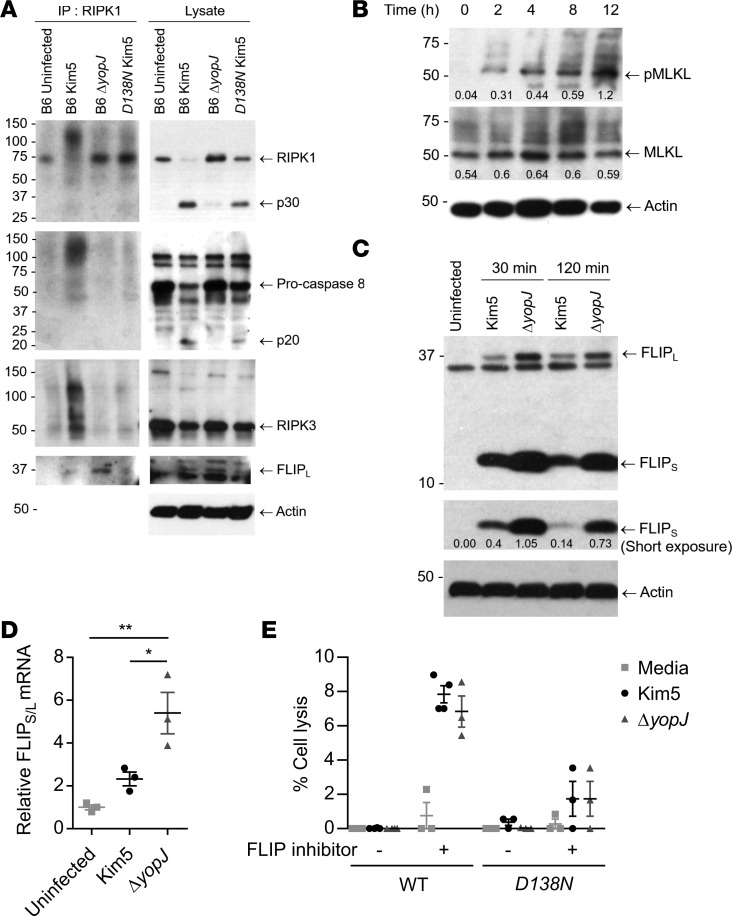
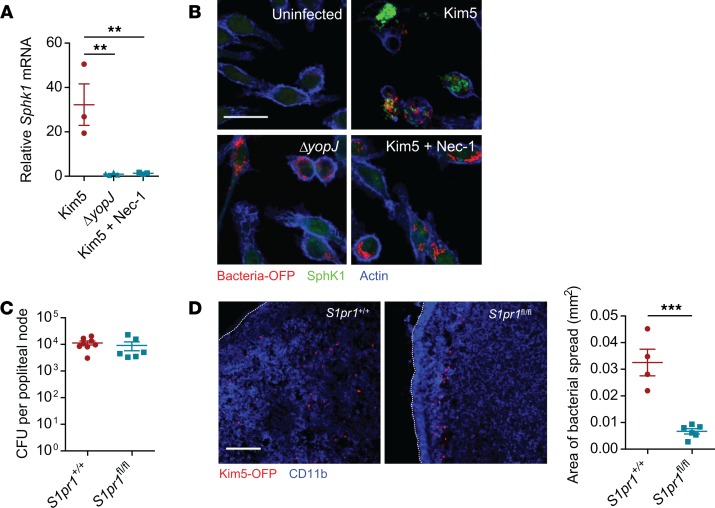
When draining lymph nodes become infected by Yersinia pestis (Y. pestis), a massive influx of phagocytic cells occurs, resulting in distended and necrotic structures known as buboes. The bubonic stage of the Y. pestis life cycle precedes septicemia, which is facilitated by trafficking of infected mononuclear phagocytes through these buboes. However, how Y. pestis convert these immunocytes recruited by host to contain the pathogen into vehicles for bacterial dispersal and the role of immune cell death in this context are unknown. We show that the lymphatic spread requires Yersinia outer protein J (YopJ), which triggers death of infected macrophages by downregulating a suppressor of receptor-interacting protein kinase 1-mediated (RIPK1-mediated) cell death programs. The YopJ-triggered cell death was identified as necroptotic, which released intracellular bacteria, allowing them to infect new neighboring cell targets. Dying macrophages also produced chemotactic sphingosine 1-phosphate, enhancing cell-to-cell contact, further promoting infection. This necroptosis-driven expansion of infected macrophages in buboes maximized the number of bacteria-bearing macrophages reaching secondary lymph nodes, leading to sepsis. In support, necrostatins confined bacteria within macrophages and protected mice from lethal infection. These findings define necrotization of buboes as a mechanism for bacterial spread and a potential target for therapeutic intervention.
Keywords: Bacterial infections; Immunology; Macrophages; Microbiology.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
